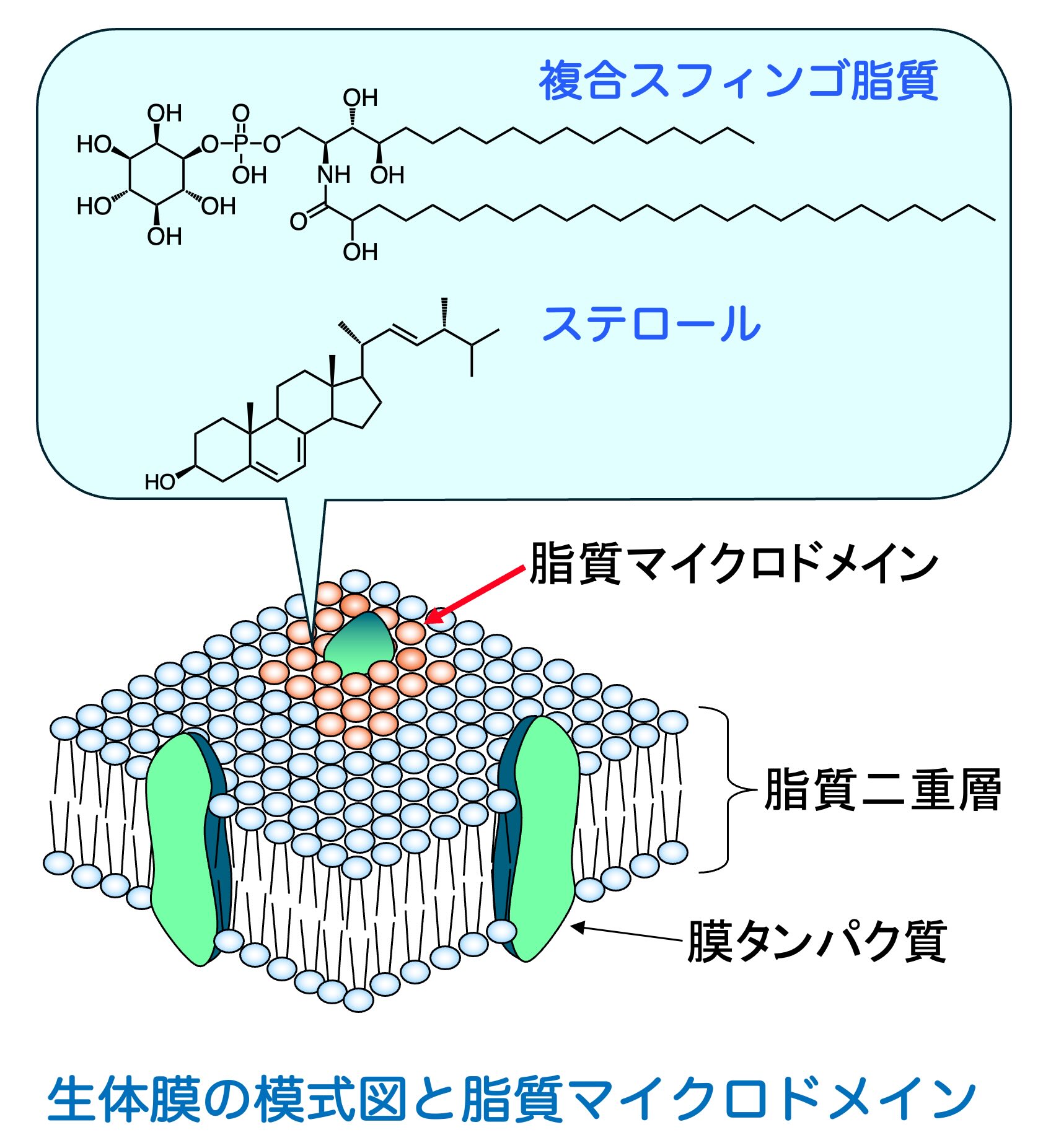
真核生物の生体膜における複合脂質には、グリセロリン脂質と複合スフィンゴ脂質が存在します。複合スフィンゴ脂質は、親水性頭部を持つスフィンゴ脂質の総称です。複合スフィンゴ脂質は生体膜上で脂質マイクロドメインを形成することで、様々な膜タンパク質が適切にはたらけるための足場を提供していると考えられています。また、その合成中間産物、分解代謝産物であるセラミド、スフィンゴイド塩基、スフィンゴイド塩基1-リン酸等が細胞内外のシグナル伝達系を制御する脂質シグナリング分子として機能します。
私たちは、「スフィンクスの謎」とも例えられるスフィンゴ脂質の生理機能を、多岐にわたる分子遺伝学的アプローチが可能な出芽酵母を用いて明らかにしようとしています。
複合スフィンゴ脂質は、膜の構造維持に寄与するだけでなく、細胞表層で進行するさまざまな生命現象を支える重要な因子です。私たちは、こうした膜脂質の構造や量の変化が、細胞の増殖、環境応答、膜タンパク質の機能発現にどのような影響を与えるかを解析しています。
特に出芽酵母をモデルとして、スフィンゴ脂質の生合成系や代謝系を分子遺伝学的に改変し、その結果として現れる表現型を詳細に比較することで、スフィンゴ脂質の多様性が細胞機能に果たす本質的な役割の解明を目指しています。
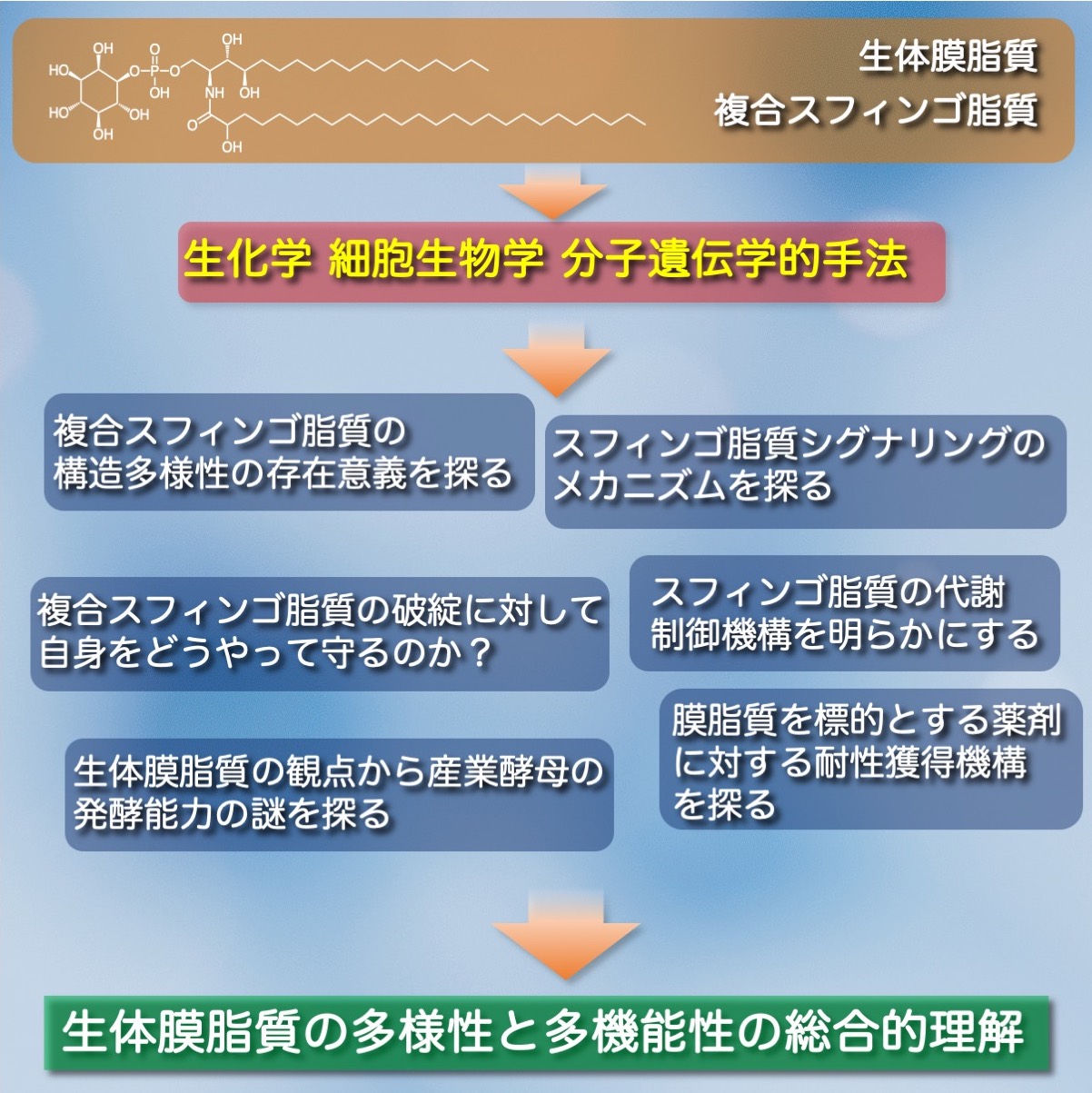
1. 複合スフィンゴ脂質の構造多様性の生物学的意義の解明

複合スフィンゴ脂質は膨大な構造多様性を持ち、これによって多彩な生理機能を発揮すると考えられています。複合スフィンゴ脂質は、一つの生物に多彩な構造バリエーションが存在するだけでなく、異種生物間においてその基本構造が全く異なる場合もあります。つまり生物界における複合スフィンゴ脂質の構造多様性の存在意義を知るためには、「一生物内における構造多様性」と「異種生物間における構造多様性」の両方の意味を理解することが大変重要です。私たちは、この両方の構造多様性の生物学的意義を出芽酵母の分子遺伝学を駆使して研究しています。
【文献】 Int J Mol Sci. 2024; 25: 12422; Trends Glycosci Glycotech. 2026; 38: E12-E16. Trends Glycosci Glycotech. 2016; 28: E109-E116.
1-1. 複合スフィンゴ脂質の構造多様性を限定した酵母の表現型解析

出芽酵母に15種類存在する複合スフィンゴ脂質の構造パターンを段階的に減らしていった変異株ライブラリーを構築することで、複合スフィンゴ脂質の多様性が限定されればされるほど種々の環境ストレスに対する耐性能が失われていくことをこれまでに明らかにしました。この耐性能の喪失は、細胞壁や形質膜のインテグリティーの損傷に起因することが示されています。現在は複合スフィンゴ脂質の多様性が、ストレス耐性能の獲得にどのようなメカニズムで寄与するのか、その分子機構の解明を試みています。
【文献】Mol Biol Cell 2022; 33: ar105; FEBS J. 2025; 292:4694-4719; Mol Microbiol. 2015; 95(4): 706-722.
1-2. 特定の複合スフィンゴ脂質分子種と他の膜脂質との機能的相互作用
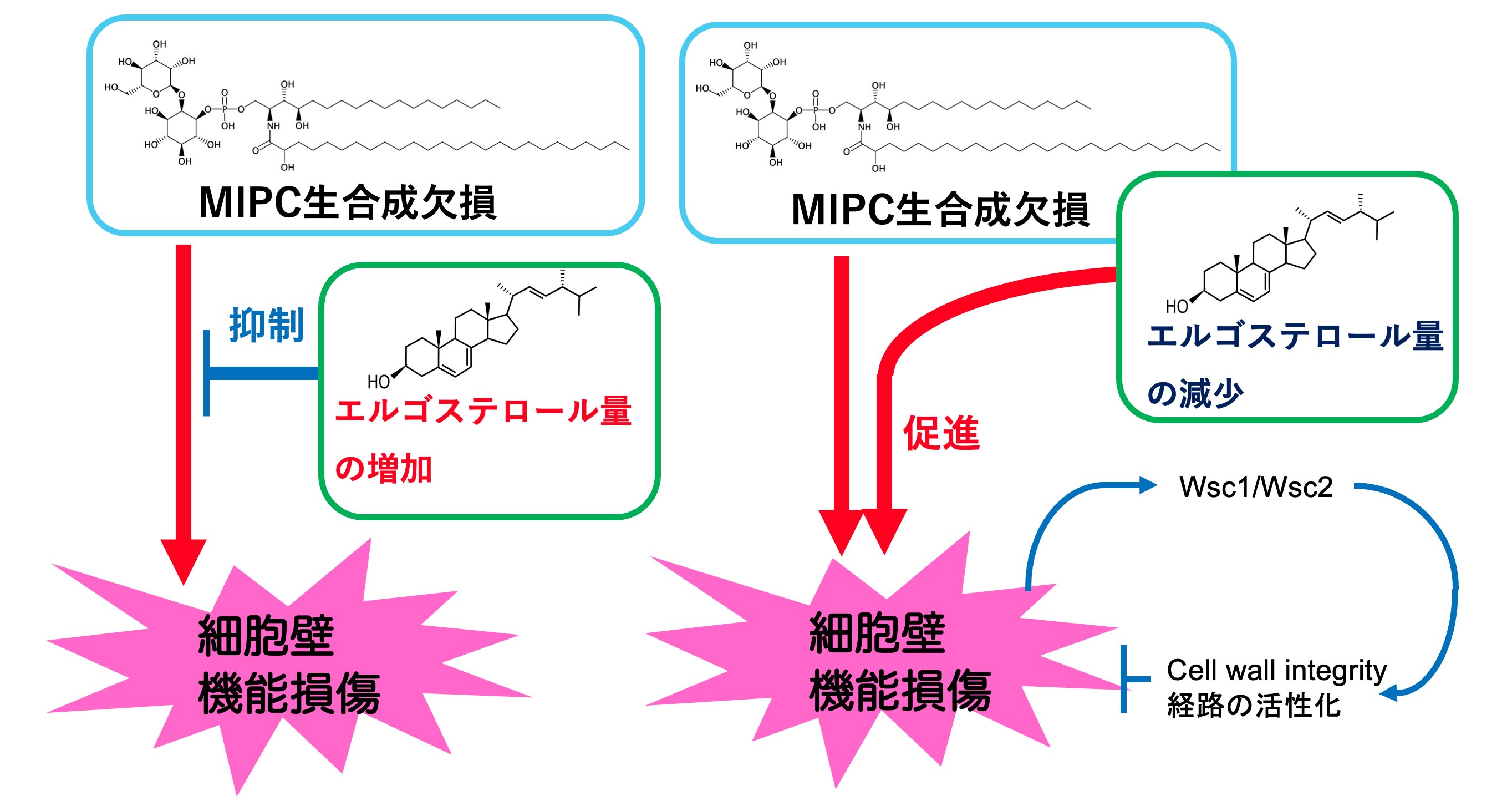
出芽酵母の複合スフィンゴ脂質の一つであるMIPCが生合成できない酵母は、細胞壁の機能損傷を示すことをこれまでに明らかにしました。さらに、エルゴステロール(酵母の主要ステロール構造)の生合成促進がこの異常を補填することも見出しました。このことは、細胞壁機能維持に二つの異なるクラスの膜脂質が協調的に関与することを示しています。またMIPCは、ホスファチジルセリンやホスホイノチシドと協調的に機能することで、エンドソームを介した小胞輸送を制御することも発見しています。
【文献】FEBS J. 2018; 285: 2405-2427; Mol Microbiol. 2012; 86(5): 1262-1280; Mol Microbiol. 2010; 78: 395-413; Biosci Biotechnol Biochem. 2020; 84: 2529-2532; Yeast. 2014; 31: 145-158.
1-3. 異種生物型のスフィンゴ脂質を持った酵母の樹立と表現型解析
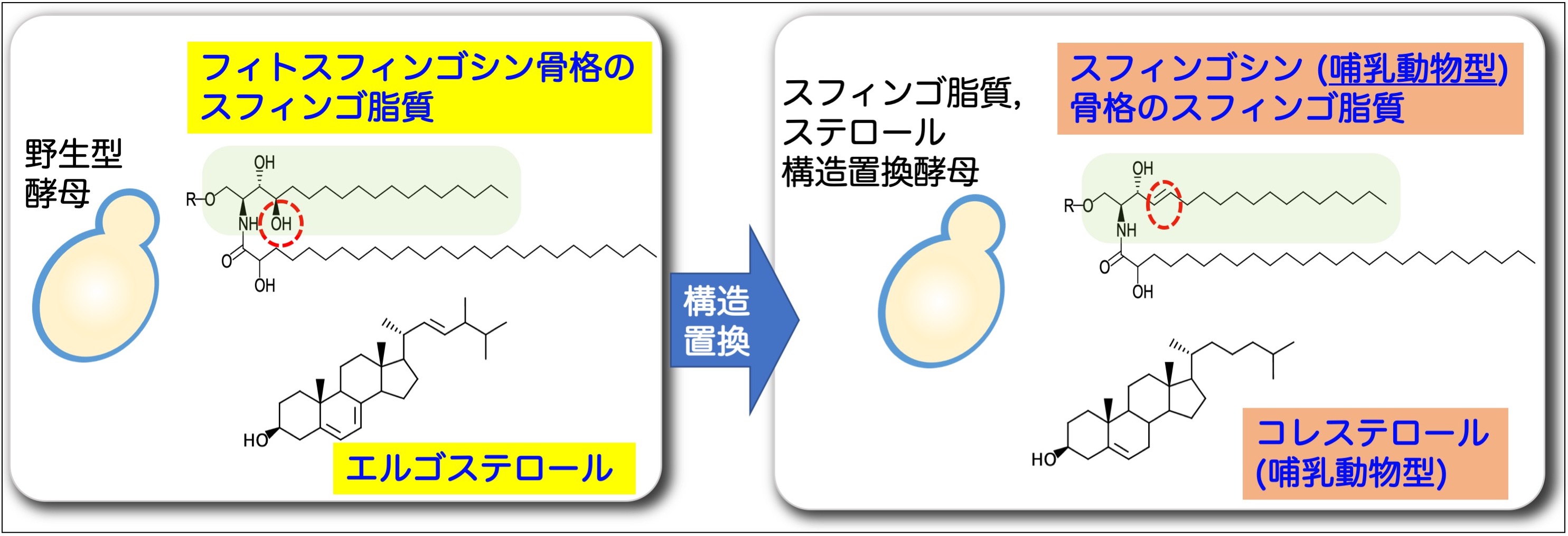
出芽酵母の複合スフィンゴ脂質の構造を異種生物のものに構造置換することで、どのような異常表現型が見られるのかを解析しています。これまでに、複合スフィンゴ脂質のスフィンゴイド塩基の生物種特異的な構造が、形質膜及び細胞壁のインテグリティーの維持に重要であることがわかってきています。このような研究を通して「なぜ、異種生物間において膜脂質の構造が異なるのか?」という基本的疑問の解明に繋げることを目指しています。
【文献】Biochem J. 2006; 394: 237-242; FEBS J. 2023; 290: 5605-5627; FEBS Open Bio 2025.
2. 複合スフィンゴ脂質代謝破綻から細胞を守る救済機構の解明
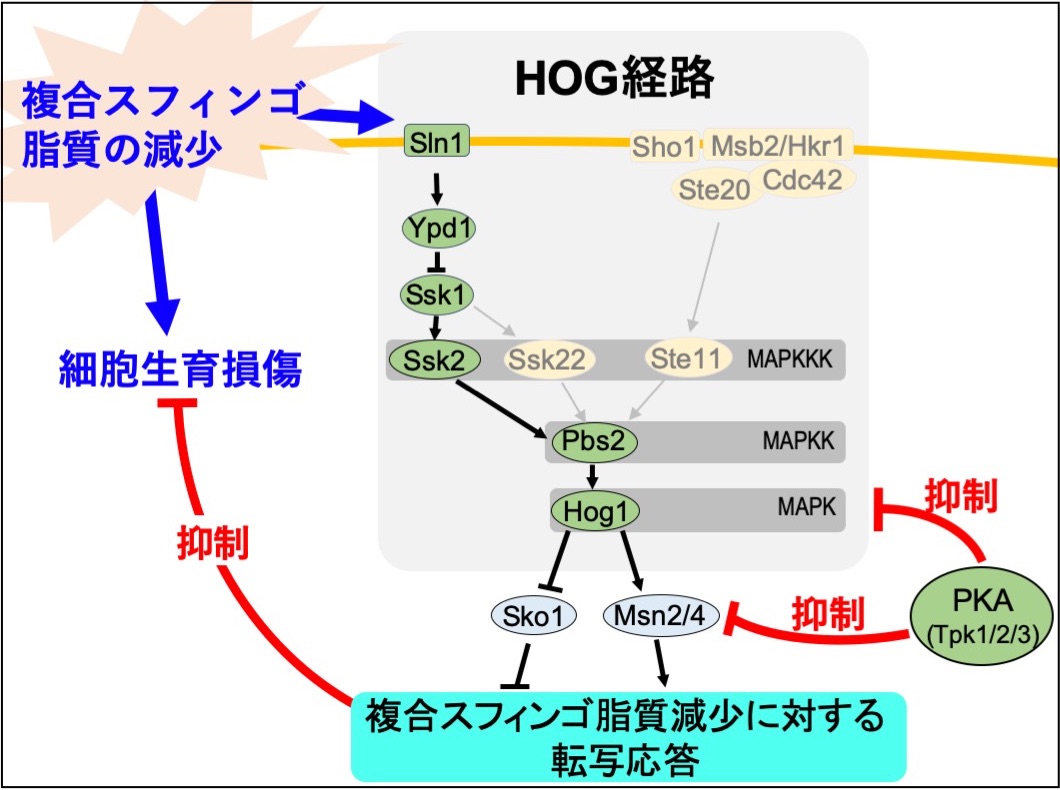
複合スフィンゴ脂質が生合成できなくなると酵母は正常な生育を維持できなくなります。私たちは、酵母の遺伝子スクリーニングによって、複合スフィンゴ脂質の生合成系が破綻した際にHOG経路と呼ばれるシグナル伝達経路が酵母の生存率を上げることを見出しています。この救済機構の分子メカニズムを明らかにすることで、複合スフィンゴ脂質の真の存在意義の解明やスフィンゴ脂質代謝異常に起因する疾患の治療戦略に繋がることを期待しています。
【文献】Mol Microbiol. 2018; 107: 363-386; FEBS J. 2022; 289: 766-786; Curr Genet. 2018; 64: 1021-1028; Mol Genet Genomics. 2026; 301: 32.
3. スフィンゴ脂質シグナリング分子の作用メカニズムの解明
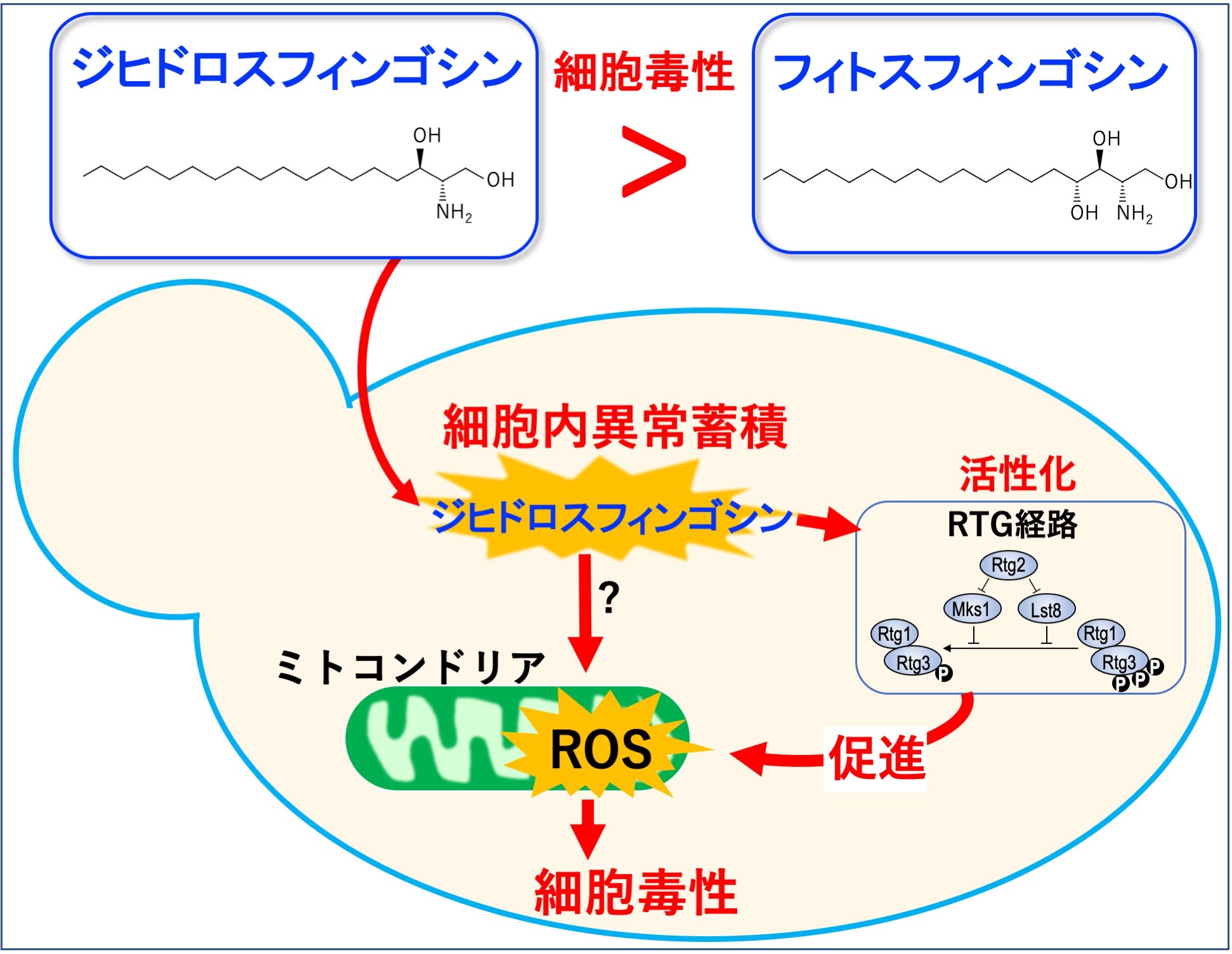
複合スフィンゴ脂質の合成中間産物、分解代謝産物であるセラミドやスフィンゴイド塩基は、脂質シグナリング分子として細胞の運命を決定する重要な分子であると考えられています。私たちは、スフィンゴイド塩基の一つであるジヒドロスフィンゴシンが酵母において、フィトスフィンゴシン(酵母の主要なスフィンゴイド塩基構造)よりも強い細胞死誘導活性を持つことを明らかにしています。また、セラミドの生理活性と拮抗的にはたらくシグナル伝達系の存在も示しています。
【文献】FEBS J. 2020; 287: 3427-3448; Biochem Biophys Res Commun. 2022; 605: 63-69; Biochem Biophys Res Commun. 2012; 417(2): 673-678; J Biochem. 2010; 148: 565-571.
4. スフィンゴ脂質代謝調節による危機回避機構
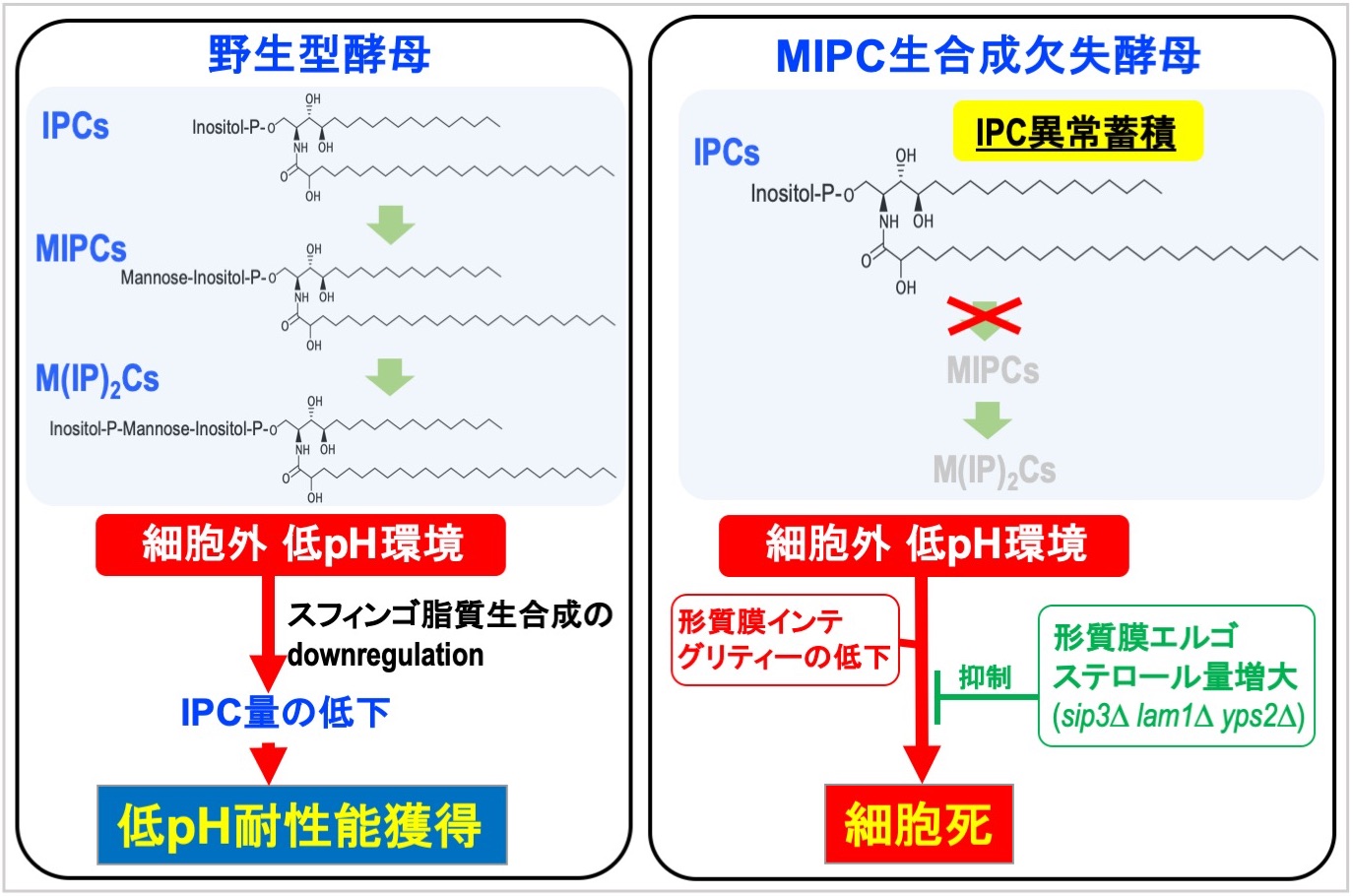
スフィンゴ脂質は様々な細胞機能の調節に関わる多機能性脂質です。私たちは、細胞内外のpHの急激な変化、もしくはエンドサイトーシスの異常下において、酵母が能動的にスフィンゴ脂質の量や組成パターンを変化(IPC低下、MIPC増大、スフィンゴ脂質総量の変化等)させることで危機回避を行なっていることを見出しました。このことは、酵母はスフィンゴ脂質の代謝を巧みに制御することで、自身の生育を脅かす様々な局面に対する防御応答をしていることを示しています。
【文献】Sci Rep. 2020; 10: 10792; FEBS J. 2016; 283: 2911-2928; Microbiology-Sgm 2015; 161: 2369-2383; FEMS Microbiol Lett. 2014; 358(1): 64-71.
5. スフィンゴ脂質を標的とした抗生物質に対する耐性獲得機構の解明
抗生物質とは特定の微生物の増殖を抑える化合物のことを指します。一方、微生物には抗生物質に対して抵抗を示す力も備わっており、抗生物質の使用によって薬剤耐性菌が出現することは医療現場での大きな問題となっています。私たちは、スフィンゴ脂質を標的とした抗生物質に対して抵抗性を付与する新たなタンパク質を同定しており、その耐性獲得メカニズムを探求しています。
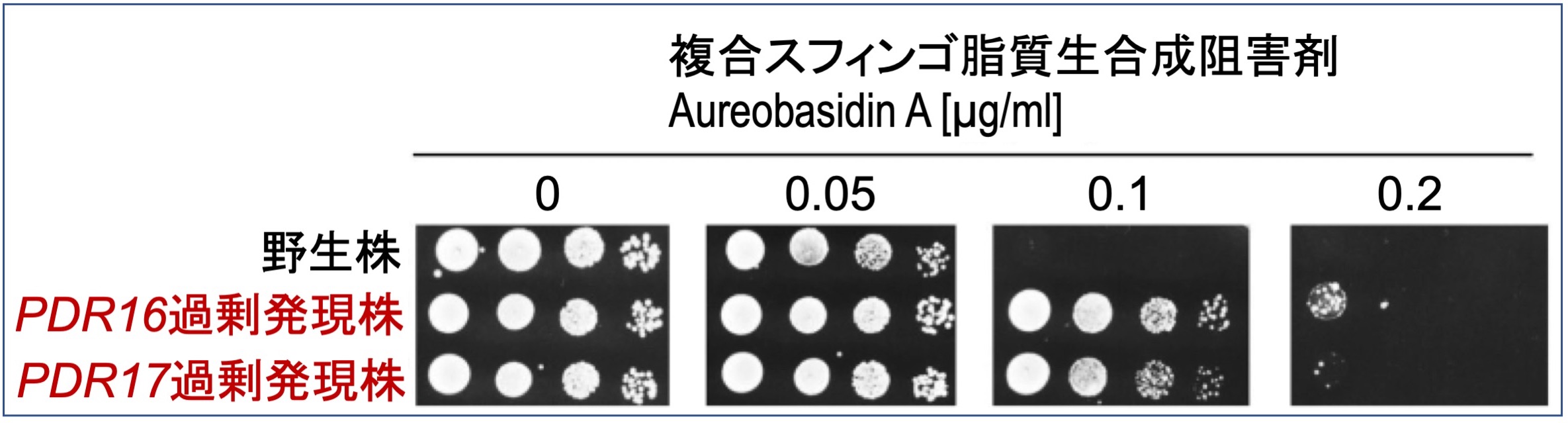
【文献】FEMS Microbiol Lett. 2018; 365: fnx255; Sci Rep. 2023; 13: 11179; Biochem Biophys Rep. 2024; 39: 101785.