![]()
植物バイオテクノロジー(植物育種学)の講義内容
目次
Ⅰ.バイオテクノロジーの定義
1.バイオテクノロジーの定義
2.植物組織細胞培養の歴史
3.園芸植物の分類
(1)学名による植物の分類
(2)植物分類の重要性
4.植物の遺伝とクローン
(1)種子繁殖と栄養繁殖
(2)ポリジ-ン(微働遺伝子)とメジャ-ジ-ン(主働遺伝子)
(3)F1ハイブリッド
(4)雄性不稔
(5)純系の作出法
(a) 同系交配または集団固定育種による純系の作出
(b) 葯培養とコルヒチン処理(半数体育種)
(6)クロ-ン(clone)
Ⅱ.植物バイオテクノロジーの基本技術
1.茎頂組織の構造
2.茎頂組織の成長と分化
(1)茎頂培養
(2)ウィルスフリー
3.不定芽分化
4.不定根形成
Ⅲ.植物の増殖技術
1.カルスと不定芽形成
2.多芽体形成(茎頂培養からの大量増殖)
(a)早生分枝
(b)多芽体
3.シュートの発生と節培養
(a)節培養
4.不定根形成
Ⅳ.順化と鉢上げ
Ⅳ.貯蔵器官の増殖
1.マイクロチューバー
2.不定胚形成
3.人工種子
Ⅴ.培養組織の長期保存
Ⅵ.植物の育種技術
1.プロトプラスト培養と細胞融合
2.細胞選抜
3.遺伝子組み換え
Ⅶ.知的財産権としての植物育成者権
Ⅰ.バイオテクノロジーの定義
1.バイオテクノロジーの定義
植物の組織培養や細胞培養は,現在バイオテクノロジーの1分野として位置づけられており,注目されている。バイオテクノロジーの定義として,1982年にベルギーで開催された国際純正応用化学連合(IUPAC)総会で以下の様な定義がなされた。
"include the application of biochemistry, biology, microbiology and chemical engineering to industrial processes and products including here the products in health care, energy, and agriculture and on the environment"
植物のバイオテクノロジーの定義をこれにしたがって行うと,「植物の生体及びその機能の操作とそれに関連するエンジニアリングの総称であり,産業的応用を目的とする」と言うことになり,実際には種苗生産,有用代謝物質の大量生産,新種苗の育種などが挙げられる。
 第1図
第1図
近年のバイオテクノロジーの中の組織細胞培養の分野についてみると
バイオテクノロジーに関係のある分野
医療――――――――がん,遺伝子病,難病治療
医薬――――――――インターフェロン,生長ホルモンの製造
環境――――――――汚染の浄化,汚物処理
バイオマス―――――人工飼料生産,バイオエタノール
食品――――――――醸造(バイオリアクター),有効成分の大量生産
水産――――――――雌性発生,三倍体魚(第2図)
畜産――――――――卵分割,受精卵移植
農林――――――――優良種苗の品種改良とその増殖,植物工場,遺伝子組み換え
バイオテクノロジーが注目された理由は?
1.DNAの発見により生物体を物理・化学および生物学の知識をもって操作できる可能性が出てきたこと,さらにライフサイエンスとして発展してきた近年の科学の延長線上にあったことがその基盤となっている。また,これらの学問的な知識が工業および農業に応用できる可能性が生じたことも大きな要因となっていると考えられる。(分子生物学による関係を強調する。タンパク質(バイオセンサー)等も含む。)
2.農業に対する企業の考え方に大きな変化が生じ,企業がこの分野に進出するようになり,その方向として種苗に着目するようになった。その結果,種子戦争と呼ばれる動きが主として外国(アメリカ)を中心に始まり,”種子を制する者は世界を制す”という言葉に象徴されるように,農業が企業にとっても大きな市場となってきた。
3.種苗法の改訂により種子や品種が特許の対象となり,”育種産業”という言葉に代表されるように育種が企業の産業対象となってきた。
4.バイオテクノロジーの技術自体が育種・大量増殖・工業化に大きな夢を持たせるに充分な魅力あるものであった。
5.資源(エネルギーを含む)の有限性の認識により工業の限界が見え始め,バイオエタノールを始めとする植物生産物の有効利用が注目され始めた。また,生物体を用いた場合その二次産物の分解が工業生産物の廃棄物よりも処理し易い。
6.工業製品の精度の限界が推測されるようになり,生物体の機能を用いた製品を開発することにより,これを補えるようになった。(バイオコンピューターなど)
植物のバイオテクノロジーとは?
植物バイオは大きく分けて,化学分野の技術と農業分野の技術に分けられる。
すなわち化学分野のものは,製薬関連企業で行われている薬効成分の抽出に代表されるもので,試験管の中で植物組織を増殖し,それから有効成分を取り出すものである。現在,実用化されているものとしては,朝鮮ニンジンやオウレン,ムラサキなどがある。
農業分野に関連する技術は,苗の大量増殖,無病苗の作出,新品種の育成,遺伝子組み換え植物などがあり,一般に植物バイオと言えばこの農業分野の技術を指すこともある。
◎農業分野におけるバイオテクノロジーの展開
植物バイオ技術と言われるものの中に,キーテクノロジーといわれる基本的な技術があり,組織・細胞培養,細胞融合,遺伝子組換えなどがそれに相当する。
主として育種,大量増殖,遺伝資源の保存,遺伝子組み換えの4つが大きな柱となっている。
2.植物組織細胞培養の歴史
Sachs(1860),Knop(1861)は植物に必要な元素の研究を行い,Sachs液,Knop液を考案した。これによって,「植物は土がなくては生育はできない」という考えが改められるようになり,「水耕栽培の祖」といわれている。
20世紀に,ドイツの植物学者Haberlandt(1902)は植物組織をガラス容器内で培養することを試みた。しかし,この試みは失敗に終ったが,その際に組織の切断面に細胞分裂を促進する物質が分泌され,それと同時にその切り口に不定形の細胞塊が形成されることを観察し,物質を”癒傷ホルモン”,細胞塊を”カルス”と命名した。同じ頃,Goebel(1902)は細胞1個は器官を構成する最低単位であり,1個体としての機能を持っており,植物体を作る能力が備っていると考え,「植物の体細胞は,生きている限り単独状態でも適当な条件に置かれれば,最小単位の生きものとして機能し,分裂と増殖を行い1個の植物体に至るまでの発育能力を潜在的に持つ」という全能性または全形成能(totipotency)の概念を明らかにした。この考えは約60年後に実証されることとなる。
1910年代になって動物の組織培養が行われ,特にその培養液として血液や幼胚のジュ-スが用いられるようになり,植物の組織培養においても,より栄養に富んだ培地で培養を行う方向へと発展するきっかけとなった。(動物細胞と植物細胞の全能性の差について説明する。動物は全能性なし。植物は全能性あり。)
Moliard(1921),Robbins(1922)は,実生の根端組織や幼芽の先端を無機養分にペプトン,アスパラギン,グルコ-スあるいはフラクト-スを添加した培地で培養し,それらを生長させることができた。特に芽からは緑色の小葉が形成されるまでにいたった。しかし,その生長も1ヵ月にとどまり,それ以上生育をさせることはできなかった。
White(1933)はハコベの茎頂の組織をボルボックスの培養に用いられた無機塩養液を基本培地とし,これにグルコ-スと酵母抽出物を加えた培地で培養し,長期間細胞分裂を観察した。また,トマトの根端をフラスコの中で培養し,伸長した根端を切り取り新たな培地に植え換えるという方法で継続的に生長させた。この根端は28年間1600代を経ても生長続けたと言われる。彼は1936年に組織培養の定義として”分離された1細胞,または数個の体細胞が in vitro で正常な生活作用を営むとき,このような培養を組織培養という”と提唱した。このような業績から彼は植物組織培養の創始者の一人であると言われる。(単細胞生物と多細胞生物の違いについて説明。器官分化)
Gautheret(1934)は,Whiteとはまったく独自に樹木の形成層を培養し,均一な組織としての無限生長を観察している。彼は,培養組織を細密に観察し,1細胞が分裂し層状に並び分裂を繰り返すことを報告した。また,1959年には「植物組織の培養」と題して,実際の方法やそれまでに出された報告書をまとめ,集大成を行った。
微量元素の作用についても研究されるようになり,Robbins(1936)は根端培養には亜鉛,マンガン,ホウ素が必要であることを指摘した。またHeller(1953)は,さらに多くの元素についても検討を行い,Hellerの基本培地を発表した。さらに,Murashige & Skoog(1962)はタバコの培養細胞の増殖に最適な培地を考案した。
基本培地に添加する物質として,酵母抽出液が有効であることが知られるようになり,その有効成分の探求が行われるようになり,アミノ酸やビタミン類がその主役を占めていることが明らかとなった。
これらの植物組織培養とは別に,植物ホルモンの研究がBoysen(1910),Paal(1919),Went(1928)によって進められ,オ-キシンが発見され,これはすぐに組織培養にも取り入れられることとなった。酵母抽出液に次いでココナツミルク(ココヤシの胚乳液)が培養に有効であることが明かとなり,同様な効果は核酸分解物やアデニン化合物でも観察され,1960年代になってサイトカイニンが発見された。これらの2つの植物ホルモンの発見は植物組織培養において飛躍的な発展をもたらした。
Reinert(1956)はカルス組織を液体培地中で振とう培養し,単細胞を取り出しそれを培養して植物体に再生できることを明らかにした。これにより植物細胞の全能性を証明できたこととなった。この研究が大きな弾みとなりその後プロトプラストさらには細胞融合へと進んで行った。
しかし,細胞融合は複数の植物の遺伝子を1つの個体が持つことになり,例えば光合成などの基本的な生命活動に関わる生理作用の発現に対して複数の遺伝子が関与することとなるため様々な問題が生じることが判り,現実的な育種操作として有効ではないことが明らかとなり,育種手法として廃れていった。
これに対してシロイヌナズナを始めとしたDNA解析によって,植物の形質発現に関わる遺伝子が解明され始めると,その特有の遺伝子のみを導入するための手法として遺伝子組み換え技術が開発され,大きく発展した。
しかし,遺伝子組み換え作物に対する社会的評価が大きく二分され始め,社会的評価を考慮した技術開発が不可欠となった。
第3図
現在までの植物組織細胞培養の流れを大きくとらえると次のようになる。
1.1900年初期における組織培養の探索期
2.1930-1960年における培地の開発期
3.1960年以降の茎頂培養,葯培養,プロトプラスト培養に代表される培養発展期
4.1980年以降の培養手法の農業への応用期
5.1990年以降の分子生物学の発展に伴う遺伝子解析と遺伝子組み換え
6.2000年以降の遺伝子組み換え作物に対する社会的評価
3.園芸植物の分類
(1)学名による植物の分類
植物の分類に関する研究の歴史は古く,植物の命名に国際的な規則を作り,世界中で共通した名前で表記するようにしている。植物分類学では表記をラテン語で行う。
植物の自然分類は,植物分類学に基づく分類で,国際植物命名規約によって規定されており,門(Division) 綱(Class) 目(Order) 科(Family) 族(Tribe) 属(Genus) 節(Section) 種(Species) 変種(Variety) 品種(Form)となっている。
一般に園芸植物の命名にはリンネが提唱した二命法が用いられており,属名と種名が併記される。学名に用いられるラテン語はローマ帝国の公用語として広く普及し,帝国滅亡後も西ヨーロッパを中心に広く使われ多言語で,民族・国境を超えて共通的に理解できる言語として現在でも専門用語や学術用語などに用いられている。
例えば,日本国内ではコメやイネと言えば日本国民は当然理解できるが,中国では理解できません。「米」や「稲」は日中共通ではあるものの,英語圏では理解できません。英語のriceは世界共通のように見えるが,アラビア語圏では全く判らないものとなり,世界中で共通した言語とはなり得ません。しかし,ラテン語のOryza sativaは民族や国に関わりなく民族感情を超えて利用できることから,極めて有利な表記法といえる。
Oryzaは属名であり,sativaは種名である。sativaの種名は「栽培された」という意味のラテン語に由来する。日本米とタイ米は食味が大きく異なるが,その学名は日本米がジャポニカ(ヤポニカ)種 Oryza sativa subsp. japonica(日本型) であり,太米の学名はインディカ種 Oryza sativa subsp. indica (インド型)となっている。日本米,タイ米共に二命法ではOryza sativaであり,亜種(subspecies)の段階で異なっている。従って,両種は極めて近縁な植物であることが理解できる。(第4図)
これに対して,西アフリカを中心に栽培されているアフリカ米の学名は Oryza glaberrima(アフリカイネ・グラベリマ)はOryza属は同じであるが,種名が異なることからOryza sativaとは植物分類学的には離れた植物であることが判る。とは言ってもコムギの学名Triticum aestivumと比べると属名が異なることから,比較的日本米とアフリカ米は近い関係にあると判断することができる。
このように,イネの学名がOryzaだと知っていれば,Oryza glaberrimaという植物名を聞いて,すぐにイネと近縁な植物だと判断でき,これは学名が自然分類に基づいて名付けられており,近縁関係を示す性格を兼ね備えているからである。
このように属名は植物グループの特性を大まかに表現できることから,園芸植物,特に花き植物では属名(学名)を一般に用いる場合が多い。
例えば園芸名サルビアの和名はヒゴロモソウ(緋衣草)で,この名前はSalvia属に属する植物の中で園芸的に利用されているものの総称である。しかし,現在ではヒゴロモソウの名前はほとんど使用されておらず,属名のサルビアが用いられており,最も一般的なものはSalvia splendensで,Salvia splendens ‘Hot Jazz’,Salvia splendens ‘Torchlight’などがある。(第5図)
サルビア属は花壇苗として様々な種が用いられており,Salvia coccinea ‘Brenthurst’,Salvia elegans ‘Honeymelon’,Salvia guaranitica ‘Omahagold’,Salvia microphylla ‘Caramba’,Salvia reptans ‘West Texas Form’などがある。
(2)植物分類の重要性
学名で用いられる属や種は植物の近縁関係を表している。種の下位の亜種が異なるOryza sativa subsp. japonicaとOryza sativa subsp. indicaは極めて近縁であり,両種は交雑が可能である。これに対して種名が異なるOryza sativaとOryza glaberrimaは普通の状態では交配することが難しく,種間交雑種を得るためには組織培養の手法を用いる必要がある。
Salvia属と近縁の属であるMosla属(イヌコウジュ属)には日本自生のMosla dianthera(ヒメジソ)があるが,これとSalvia coccineaとの間はかなり離れており,属間雑種を育成しようとする場合には極めて高度な培養技術を必要する場合が多い。
このように種が同じ植物は育種が極めて容易である。これに対して属が同じで種が異なる場合には,共通した生理機構を持っているが形質も大きく異なり,種間雑種を育成する場合には困難が生ずる場合が多い。しかし,耐病性や耐寒性などの形質を導入するためには有効な交配手法となる場合が多い。
4.植物の遺伝とクローン
(1)種子繁殖と栄養繁殖
種子繁殖=自家採種:F1ハイブリッドが出てくる以前の種子繁殖性の作物は,集団固定育種法と呼ばれる方法で育種され,この集団固定育種法は優良な形質を持つ個体を次々と選択しながら優良形質を固定する方法で,この操作を繰り返すことによってホモ個体の集団を形成することができる。
ホモ;homozygosis 同型接合;ある形質を支配する遺伝子座について対立遺伝子が同型であることをいう。対立語は,ヘテロ(heterozygosis) 。
このようにして形成された優良形質を持つ種子繁殖性の野菜や花は,自家採種によって栽培することができ,この系統は自殖によって維持されていた。
栄養繁殖;種子繁殖(有性繁殖)に対し,栄養器官の一部を切り離すか,人為的な操作を加えて不定根,不定芽などを形成させて,独立した個体を得る方法で,無性繁殖ともいう。従来の方法には,挿し木(枝挿し,葉挿し,根挿し),接ぎ木,取り木,株分け(分球を含む)などがある。これに加えて,組織培養による繁殖(micropropagation)が近年大きな栄養繁殖法として着目されており,これらの栄養繁殖法によって繁殖された個体は,突然変異が生じない科ぎり,遺伝形質が受け継がれ,種子繁殖したものに比べ開花,結実が早まることがある(実生では桃栗3年,柿8年といわれるが,接ぎ木をすると2~3年で開花する。)。また,枝変わりや雌雄株を選んで増殖することも可能なため,遺伝的に複雑な雑種性を持つ園芸植物では,主要な繁殖法となっている。
(2)ポリジ-ン(微働遺伝子)とメジャ-ジ-ン(主働遺伝子)
1遺伝子では働きが小さいが,多数の同じ遺伝子が補足しあって量的形質の発現に関わっている遺伝子を指す。これに対し,メンデル分離を示し,その働きが明確に区別できる遺伝子を主働遺伝子と言う。
(3)F1ハイブリッド
ヘテロシス;他家受精植物を自殖や近親交配を続けると生育が著しく劣る自殖弱勢を生じる場合があるが,これに同様な近交系や純系を交配すると,そのF1雑種が両親をしのぐ生育を示す場合がある。この現象をヘテロシスと言う(第6図)。これは,交雑によって遺伝子型がヘテロ接合になり,両親と異なる表現型となることをいい,雑種強勢とも言う。この現象が生じる原因として,超優勢説と優性遺伝子連鎖説がある。前者では,それぞれの対立遺伝子がヘテロ接合になることにより,優性ホモの場合より形質値が大きくなることがヘテロシスの原因であるとし,後者ではヘテロ接合が優勢ホモの表現型と同じとなるが,劣性ホモの対立遺伝子がないため,結果として形質値が大きな値を示すことになるとする説である。
トウモロコシなどの穀物では,表現形質の数量的増加(収量,葉面積の増加など)を目標として,F1ハイブリッドを用いる場合が多いが,野菜や花では,均一化や優良な形質の集積を目標としてF1ハイブリッドを用いることが多い。
(4)雄性不稔
F1ハイブリッドを用いた育種を行なう場合,必ず交配を行なう必要があり,自殖を行なわないようにする必要がある。そのためには,除雄と呼ばれる雄蕊を取り除く操作を行う必要がある。しかし,この操作は極めて多大な労力を必要とするため,花粉の発芽能力を持たないようにする遺伝子(雄性不稔遺伝子)を用いてF1ハイブリッドを作ることが行なわれている。雄性不稔性の遺伝には,核支配のもの,細胞質支配のもの,核と細胞質の両者の支配によるものの3種類が知られているが,組織細胞培養で有効なものはこの中で細胞質遺伝子由来の雄性不稔である。細胞質由来の遺伝子には大きく分けて2種類あり,ミトコンドリアDNAと葉緑体DNAが挙げられる(ミトコンドリアと葉緑体にDNAがある理由)。細胞融合による雄性不稔系の作出と交配による雄性不稔系の作出の説明をする。
(5)純系の作出法
(a) 同系交配または集団固定育種による純系の作出
種子繁殖性の品種の維持は,特定の形質に関して純系を維持する必要がある。自殖を継続して行うことで純系を作出することは可能であるが,様々な形質に関してホモ(homozygosis 同型接合)となることによって,劣性形質の蓄積が行われ始め,自殖弱勢と呼ばれる現象が生ずる。これを避けるために,特定の形質(例えば,イネでは着粒数や粒径,成分など)だけに着目し,これ以外の形質に関しては選抜淘汰圧をかけないで交雑を行う。
得られた個体の中からさらに特定の形質について優良な個体を選抜・交雑を繰り返すことで,自殖弱勢を発現させることなく特定の形質に関して純系の個体を選抜することが可能となる。
イネなどの穀物では,純系の作出法として同系交配や集団固定育種が用いられる。
(b) 葯培養とコルヒチン処理(半数体育種)
同系交配や集団固定育種での純系の作出は多大な年月を要するため,より効率的な純系作出法として半数体育種法が用いられる。特に1回の交配で多数の種子が得られやすい果菜類などで行われている。
花粉は半数の染色体を持つが,これを組織培養の手法を用いて不定芽あるいは不定胚を形成させて植物体を得る。得られた植物体にコルヒチンなどで倍数化処理を行うと,同型接合子を持つ純系の個体を得ることができる。
(6)クロ-ン(clone)
突然変異育種は,種々の誘発源により人為的に有用な突然変異を誘発し,これを品種の形質改変に利用する方法である。
人為突然変異育種の研究は,1930年にX線を大麦とトウモロコシに照射した報告がある。その後,放射線,中性子,重イオンビームなどの照射による誘発の他,化学物質等の処理などが行われている。(第7図,第8図)
これらの突然変異個体は交雑育種を行うことでその形質発現が失われることが多く,突然変異個体が選りすぐれた特性を持つ場合には品種として登録され,組織培養などの栄養繁殖によって維持・繁殖される。
Ⅱ.植物バイオテクノロジーの基本技術
1.茎頂組織の構造
シュート(shoot)は苗条とも言い、茎と葉とからなる1つの単位を示す。芽はシュートには必ず含まれ、頂芽(apical bud)と側芽(lateral bud)がある。種子植物の側芽は必ず葉の付け根(葉腋)に存在するため、腋芽(axillary bud)とも言う。側芽は葉腋に形成され、それらの形成される部位は規則性があり、葉の配列様式(葉序)と関係がある。このように植物体の決まった位置に形成される頂芽と側芽は定芽(definite bud)と呼び、これに対し通常は芽が形成されない葉、根、カルスなどに形成された芽を不定芽(adventitious bud)という。
形成された芽は、頂端分裂組織(shoot apical meristem)を持ち、この頂端分裂組織が将来葉となる組織である葉原基(leaf primordia)を分化する。この葉原基を分化していく能力を持つ組織を茎頂(shoot apex)という。茎頂部は2層からなる外衣とその内側の内体と言われる細胞群から構成され、最も外側の外衣層は表皮組織に分化する。葉原基が形成される場合には外衣の内側の層が2層に並層分裂し、形成され始める。
茎頂組織の遺伝的安定性は高く、茎頂組織から形成されたシュートは発芽時の形態を極めて良く維持している。
2.茎頂組織の成長と分化
茎頂組織の分裂は、主にサイトカイニンによって促進される。
茎頂組織の遺伝的安定性
(1)茎頂培養(メリクロン)
頂芽あるいは側芽から1㎜程度の茎頂組織を切り出し、培地に植え付ける。培地に添加する植物ホルモンは、木本植物の場合にはサイトカイニン単独あるいはサイトカイニンとジベレリンを同時に添加する場合が多く、草本植物の場合にはサイトカイニンとオーキシンを添加する。
生長点を切り出し培養すると、生長すると共に多くの芽(腋芽)が形成され、短期間に大量の植物体を再生する。この植物(苗)を試験管から移植することによって、良く生育が揃った苗を供給することができる。また、種子では繁殖できない植物も栽培対象とすることができる。
従来の株分けや挿し木繁殖では繁殖効率が悪く、年間数十万本の植物を増殖することは困難であり、組織培養を用いた大量増殖法が花の分野において一般的技術となっている。
(2)ウィルスフリー
植物に感染するウィルスは 600種以上が知られており、これらのウィルスは感染した植物体内で維管束を経由したり細胞間を移行したりして広がり、植物体全体に分布するようになり、全身感染を引き起こす。植物は動物とは異なり、坑体による免疫機構を持たないため、いったん感染すると枯れるまでウィルスが除去されることはなく、植物のウィルス病は治癒しない。
挿し木、接ぎ木、分球などの栄養繁殖法で増殖されることの多い園芸植物では、ウィルス病は極めて重大な病気であり、従来ではウィルスに罹病した個体は廃棄処分をするより方法がなく、中世にはチューリップでウィルス病にかかった個体に出る縞状のモザイク模様を新たな品種として登録され、きわめて高価な価格で取り引きされた経緯もある。
ウィルスの植物体内の分布は一様ではなく、古い組織ほど多く、新しい組織では分布が少ない。
ウィルスは細胞壁を能動的に貫通して細胞内に進入することはできず、その感染経路は、1.植物体に機械的にできた傷や、2.昆虫、ダニ、線虫、菌類などの食害や寄生の際にできる傷から進入したり、3.接ぎ木をしたり、病花粉によって受精したりして伝播される。昆虫による感染の場合には、アブラムシ、ウンカ、ヨコバイなどの吸汁性昆虫の口針進入によってできた傷から樹液と共にウィルスを吸汁し、口針に付着したウィルスを健全植物から吸汁する際にダ液と共に吐き出させるためこれによって感染する。すべてのウィルスがこれらの方法すべてで感染するものではなく、ウィルスによって感染方法が決まっている。
植物体内の移動については、ウィルスが細胞壁を能動的に貫通できないため、細胞壁内にある原形質連絡を通じて細胞間を移動すると考えられている。また、細胞分裂によって細胞と共に広がる。さらに、師部を経由して遠隔部位に移動する。
ウィルスは茎頂組織にはほとんど分布していないことが観察されている。この理由として、1.生長点近傍組織の伸長生長がウィルスの細胞間移動速度より大きいため、ウィルスが生長点に到達できないことや、2.生長点の近傍組織が組織構造的にウィルスの進入を阻害していること、3.生長点近傍組織が生理的にウィルスが増殖できない状態におかれていることなどが考えられた。しかし、ウィルスの細胞間移動速度は生長点の生長速度より早く、1.の理由は考えられず、2.については組織構造学的に生長点の細胞とそれ以下の細胞に差がなく、否定された。古い組織に比べ新しい組織ではウィルスに対する感受性が低く、ウィルスの細胞内増殖速度も低いことが知られていることから、現在では3.の理由が茎頂組織内にウィルスが含まれない理由と考えられている。
この機構としては、以下の3点が考えられている。
1.細胞内代謝の活性が低い細胞に比べて、高い活性状態にある宿主細胞の代謝物をウィルスが摂取する事はより困難なことであると考えられる。したがって、頂端分裂組織の細胞中ではウィルスの増殖に必要なRNA合成は抑制されるものと思われる。
2.植物体にウィルス不活性系が存在するものと仮定すると、茎頂部分は他の組織部位よりもその活性が高く、このことが頂端分裂組織中の細胞をウィルス感染から保護しているのであろう。
3.培地中に添加したオーキシンはウィルスの増殖を阻害することが知られている。茎頂においてオーキシンレベルが高いことからこの内生オーキシンがウィルスの活動に抑制的に作用している可能性がある。
このようなことから、茎頂培養を行うことによってウィルスフリー植物を作出することができると考えられた。このことを実際的に考えたのはトマトの根の培養で有名な Whiteで、この事実の基づき Morel and Martin (1955)はダリアとジャガイモで実証した。
イチゴに感染するウィルスは、イチゴモットルウィルス(strawberry mottle virus;SMoV),イチゴベインバンディングウィルス(strawberry vein banding virus;SVBV),イチゴマイルドイエローエッヂウィルス(strawberry mild yellow edge virus;SMYEV),イチゴクリンクウィルス(strawbery crinkle virus;SCrV)の4種類である。これらのウィルスはアブラムシや接木によって感染する。
イチゴウィルス病の病原ウィルスと病徴
イチゴモットルウィルス(SMoV)は、単独では無病徴、しかし古い葉の枯れ込みや、ランナーの発生が少ないなど年間を通してみると生育はやや劣る。
イチゴベインバンディングウィルス(SVBV)は、単独では無病徴、SMoVと重複感染すると草勢低下。SMoVおよびSMYEVと三重感染すると発育不良、葉先のねじれ、小葉杯状がみられ、葉緑が黄化する場合もある。
イチゴマイルドイエローエッヂウィルス(SMYEV)は、単独ではほとんど無病だが、移植後や収穫後などに古い葉が突然紅葉する事がある。SMoVとの重複感染で草勢やランナー発生の低下がみられ、古い葉が枯れ易くなる。
イチゴクリンクウィルス(SCrV)は、生育にほとんど影響しないが、葉が縮葉状になる傾向がある。
これらの分布状況は、SMoVが最も広く蔓延しているが、単独ではほとんど病徴を示さないことが多く、生育障害をおこしている株はSVBVやSMYEVと重複感染している場合が多い。
◎生長点の大きさとウィルスフリー化の割合
0.2-0.4mm 100%の個体がウィルスフリーとなる。
0.2-0.8mm SVBV,SCrVは100%、SMYEVは97%、SMoVは78%が除去できる。
※ウィルスフリー苗(無病苗)
現在生育している全ての植物はウィルスを保持していると言われているが、実際にはそれらのウィルスは発病することはない。しかし、植物体がなんらかの要因で生育が低下したり、弱ったりした場合、その病兆が発現し、ひどい場合には枯死してしまう。
植物体からウィルスを除去した場合、植物体の生育は極めて良くなり、著しい場合には除去前の2ー3倍の生育量を示したり、花の場合にはその色が良くなるなどの結果が得られている。
この技術はすでにイチゴやユリ,ナガイモ,ニンニクなどの野菜や花で実用化されている。この技術は現在最も現実的な植物バイオの分野の一つである。
(第9図,第10図,第11図,第12図,第13図,第14図,第15図)
3.カルスと不定芽分化
ハクランとグリーンスポット(第17図)
4.不定根形成
Ⅲ.植物の増殖技術
第16図
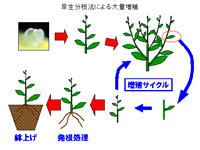
1.多芽体形成(茎頂培養からの大量増殖)
(a)早生分枝
サイトカイニンを高濃度に添加することにより、頂芽優勢が打ち消され、側芽の伸長が促進される。その結果、頂芽の伸長と同時に側芽の伸長が開始され、分枝をし続ける形態のシュートが形成される。一般に分枝は、頂芽の生長がある程度進んだ後に行われるのに対し、頂芽の生長が完全に行われないまま早期に側芽が分枝するため、早生分枝法と言われる。早生分枝形成はサイトカイニンの種類によっても差異がみられ、カキを材料とした場合においては、ゼアチンでは早生分枝が形成されず、BAPの場合には早生分枝が形成された。このサイトカイニンの種類と早生分枝形成との関係は、明らかにはなっておらず、サイトカイニンの作用である茎頂組織の分裂促進による頂芽の生長促進作用と頂芽優勢打破作用がサイトカイニンの種類によってその強弱が異なるのかもしれない。
形成されたシュート(分枝したシュート)を切り取り、新たな早生分枝形成培地に植え継ぐとこのシュートから早生分枝が形成され、増殖が図られる。
 (第28図)
(第28図)  (第20図)
(第20図)
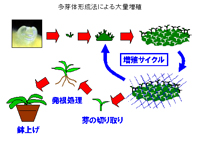
(b)多芽体
シュートの伸長をさせず、側芽を多数形成させる方法で、一般に草本で用いられる増殖法の一つである。このため培地にはサイトカイニンに加えてオーキシンを添加する。オーキシンの作用によってシュートの伸長が抑えられ、かつサイトカイニンの添加によって茎頂組織での葉数の分化が促進され、その結果多数の腋芽の形成が行われる。形成された多芽体の基部は1つにつながっているため、塊状の多数の芽を持つ組織が形成される。
これを小塊に切り分け、再び多芽体形成培地に植えることによって新たな多芽体が形成される。
 (第29図)
(第29図)
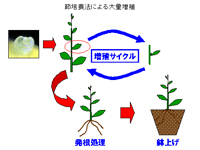
2.シュートの発生と節培養
(a)節培養
早生分枝を形成しにくい植物(頂芽優勢が強い植物)や早生分枝を形成し易いBAPなどでは生長組織の生長が促進されない植物では、ゼアチンなどを用いて一次シュートの伸長が促進される。このシュートを節毎に切り放し、その節を再び培養し、節に含まれる側芽を伸長させる培養方法で、カキはこの方法で増殖することにより1カ月半で約5倍の増殖率を維持している。
 (第30図)
(第30図)  (第18図)
(第18図)  (第19図)
(第19図)
3.不定根形成
 (第26図)
(第26図)
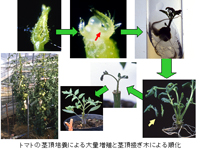
Ⅳ.順化と鉢上げ
組織培養苗が従来の苗と異なる理由の1つに、培養する瓶(フラスコ)の中の環境がよすぎるため、外の環境に耐えられないこともある。すなわち、フラスコの中は湿度が100%であり、温度も25℃で最適環境であるため、葉の表皮が発達していないことや、植物に取っては鼻に当たる気孔が発達していない。また、培地には糖分が含まれるため、本来光合成によって糖を自分で作り出す作用が低下している。
このため外気の環境に慣らせるためには、なるべく早く外気に慣れた新しい葉を出させることが必要である。
 (第21図)
(第21図)
Ⅳ.貯蔵器官の増殖
1.マイクロチューバー
貯蔵器官であるマイクロチューバーの形成には糖濃度が密接に関与している。糖濃度が高まることによって栄養成長から貯蔵器官形成への誘導が行われる。ジャガイモのマイクロチューバー形成にも登濃度が密接に関与しており、茎頂培養によって増殖したシュートを糖濃度が高い培地を添加することによって、腋芽が肥大を始めマイクロチューバーが形成される。
同様にシクラメンにおいても液体振とう培養を行うことでマイクロチューバーが増殖し、同時に肥大させることができる。
マイクロチューバーに類似した貯蔵組織の増殖事例としてユリの球根の大量増殖がある。茎頂培養の後に形成された子球をサイトカイニン濃度の高い培地で培養することによって多芽体が形成され、それを細断して糖濃度の高い培地に移植すると、多芽体に含まれる茎頂組織が肥大を始め、小球が形成される。これをマイクロチューバーと同様に定植することが可能である。
このようにして形成させたマイクロチューバーは貯蔵器官であることから、一定期間貯蔵することが可能であると同時に、多芽体形成や節培養などで増殖した培養個体のように順化処理を行うことなく定植し、発芽と同時に苗生産が可能となる。
 (第22図)
(第22図)  (第23図)
(第23図)
2.不定胚形成
マイクロチューバーと類似した貯蔵組織の1つに胚組織がある。胚は種子の中に含まれる器官で、高い貯蔵性を持つと共に容易に植物体として再生させることが可能である。
不定胚形成は全ての細胞から行われる訳ではなく、特定の形質を有するEmbryogenic Callusと呼ばれる細胞(組織)から形成される。Embryogenic Callusの分化要因には糖飢餓や重金属などのストレスが関与していると推定されており、不定胚形成を誘導する特定のタンパク質が単離されている。しかし、Embryogenic Callusの分化は必ずしも完全に制御されているとは言えない状況で、今後の研究の進展が待たれる。
 (第24図)
(第24図)
3.人工種子
不定胚はマイクロチューバーとは異なり、そのままの状態では培養容器外で発芽させることは難しい。その理由として、マイクロチューバーのように貯蔵養分を持たないことに加えて、外界に存在する病菌や環境変化に対して耐性を持たないことが挙げられる。そのために、不定胚を保護しながら発芽に必要な養分を供給するための組織(種子では種皮と胚乳に相当する組織)を提供する必要がある。
種皮と胚乳に相当するものとしてアルギン酸カルシウムなどのゲル状物質を用いたカプセルの中に不定胚を封入することによって種子と相同な人工種子を作成し、発芽させることが可能となる。(第25図)
Ⅴ.培養組織の長期保存
植物の遺伝資源の保存は,一般には栽培の継続によって行われている。しかし,植物の栽培は栽培圃場や栽培労力の確保に加えて,気候の変動あるいは栽培管理技術の問題など,確実な遺伝資源の保存を遂行することが困難となる場合がある。
一般の組織培養では培養施設の面積は小さいものの,温度や日射量などの環境制御に多大な維持管理経費を必要とし,さらに定期的な継代培養のための労力も必要とする。
組織培養技術を用いた遺伝資源の長期保存の取組としては,野菜や果樹での取組がある。サツマイモでは,培地に植物ホルモンを添加せず,さらにマニトールを用いて糖吸収を抑える成長抑制培地を用いて20ヶ月以上の培養期間で継代培養を行わずに維持保存が可能であることが報告されている。また,長期保存培養後の栽培においても変異個体の発生が認められなかった(埼玉園試1997)。
果樹などの温帯性木本植物や温帯・寒帯性植物においては,培養を5℃程度の低温庫内で行うことで成長を著しく遅延させ,かつ低温障害の発生が見られないことから,低温培養を行うことによって継代培養を行うことなく1年以上培養を継続させることができる。
また,茎頂などの微細組織を-150℃以下の液体窒素で凍結保存する取組も各種作物で行われており,数年間の長期保存が可能であることが実証されている。
突然変異によって生じた雄性不稔や種子形成能力のない1年性植物を栽培で維持することは極めて困難であるが,組織培養技術を用いることでこれらの個体を維持することも可能となっている。
Ⅵ.植物の育種技術
1.プロトプラスト培養と細胞融合
植物細胞は細胞壁を持つことから,動物細胞や微生物などの単細胞生物で行われてきた細胞融合や外来遺伝子の導入が困難であった。 1960年にCockingが酵素処理によって細胞壁を持たないプロトプラストの単離に成功し,1971年に長田・建部がプロトプラストから植物体を再生することに成功して以来,急速にプロトプラストを用いた研究が行われた。
当初のプロトプラストの研究は細胞融合を目的とした研究が主流で,単離したプロトプラストをポリエチレングリコールや電気刺激などによって融合させ、雑種細胞が作出された。細胞融合によって作出されたものとして,ポマト(トマトとジャガイモの細胞融合雑種)、オレタチ(オレンジとカラタチの細胞融合雑種)やイネとダイズの細胞融合雑種などが挙げられる。
しかし,細胞融合は異種間の細胞全体を融合するため,各々の細胞が持つ共通の遺伝子,例えば光合成や呼吸などの遺伝子を複数持つことになり,それらの遺伝子が相互に干渉しあうために正常な成長が見られないことが多く,実用的な個体は未だ得られていない。
このような状況から,プロトプラストを用いた細胞融合に関わる研究は急速に減少し,特定の遺伝子を導入する遺伝子組み換え技術へと変化した。
現在,細胞融合で期待されている分野は,同一植物のプロトプラストを細胞融合して倍数体の作出に留まっており,この他では体細胞変異の効率的な作出を目的とした細胞選抜がある。
 (第27図) (第31図)
(第27図) (第31図) 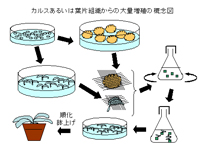 (第33図)
(第33図) 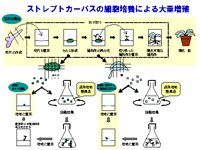 (第32図)
(第32図)
2.細胞選抜
植物の細胞は1個の受精卵の細胞分裂によって形成されるため,理論的には同一遺伝子を持つ細胞から形成されている。しかし,実際には成長点組織以外の細胞では様々な変異が生じており,この中には形質が発現されないものの有用な突然変異が起きている可能性がある。また,葉肉細胞や貯蔵組織の細胞は自然倍数性を持っているものもあり,それらの細胞を培養して植物体を再生することで倍数体の作出も可能である。主な細胞選抜の成果として,耐病性細胞からの植物体再生による耐病性個体の育成を挙げることができる。
また,カルスは特定の器官形成能力を持たない細胞塊の集合体で,高頻度の突然変異が起きているといわれている。この特性を利用して突然変異個体の選抜を行ったり,カルスに放射線を照射して人為的に突然変異を誘発させ,その後に細胞培養を行って有用な突然変異個体を選抜する試みが行われている。
細胞選抜で最も重要なことは選抜に用いるストレスを限定させることである。細胞選抜で成果を挙げているものとして,病原性毒素を培地に添加した耐病性細胞の選抜を挙げることができ,フザリウム(立枯病)やバーティシリウム(萎ちょう病),青枯病などに対する耐病性品種が育成されている。
3.遺伝子組み換え
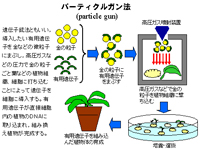
遺伝子組み換え法にはパーティクルガン法(particle gun),エレクトロポレーション法(electroporation),アグロバクテリウム法(Agrobacterium) 3種類の方法がある。パーティクルガン法は遺伝子銃法ともいい,導入したい有用遺伝子を金などの微粒子にまぶし,高圧ガスなどの圧力で金の粒子ごと葉などの植物組織、細胞に打ち込むことによって遺伝子を細胞に導入する。有用遺伝子が直接細胞内の植物のDNAに取り込まれ,組み換え植物が完成する。エレクトロポレーション法は電気穿孔法ともいい,プロトプラストと有用遺伝子を溶液に入れ,直流の電気パルス(数1000v/cmの高電圧で数10μ秒のパルス)をかけるとプロトプラストの細胞膜に短時間小さな穴があき,外液と共に遺伝子が導入される。有用遺伝子が目的の植物のDNAに取り込まれた後,プロトプラストを培養し,植物体を細分化させて組み換え植物を作る。
アグロバクテリウム法はAgrobacterium tumefaciens(アグロバクテリウム ツメファシエンス)という植物に根頭癌腫病を引き起こす土壌細菌を利用します。この土壌細菌は植物の傷口などから植物に感染し,土壌細菌が持っている環状DNAの一部分が植物の染色体上に組み込まれ,遺伝子組み換えが起こすことが1970年代にわかってきた。この研究を基に1980年代に入ると遺伝子組み換えの研究が盛んになり,1984年にはHorschにより世界で最初の遺伝子組み換え植物がタバコで作り出された。他の植物などが持っている耐病性などの有用遺伝子を土壌細菌の環状DNAに入れて植物に感染させ,その有用遺伝子が植物細胞の染色体に組み込ませ,耐病性などの新しい性質を持った遺伝子組み換え植物が得られる。
土壌細菌のAgrobacterium tumefaciensは双子葉植物にのみ感染するため,当初アグロバクテリウム法はイネやトウモロコシ,コムギなどの有用作物に適用できず,パーティクルガン法やエレクトロポレーション法が用いられていた。しかし,Agrobacterium tumefaciensに関する研究が進み,Agrobacterium tumefaciensの植物細胞への感染にアセトシリンゴン(Acetosyringone)などの物質が密接に関係し,双子葉植物ではアセトシリンゴン(Acetosyringone)などの物質を生合成できるのに対して単子葉植物ではその生合成能力がないために感染できないことが明らかとなり,単子葉植物に対しては人工的にアセトシリンゴンなどを加えることで感染させられることが明らかとなった。
現在では,遺伝子組み換え法としてアグロバクテリウム法が最も汎用的に用いられている。
最初の農作物としての遺伝子組み換え植物はトマトのフレーバーセーバー(Flavr Savr)で,1992年にCalgene社によって作られ,アメリカ食品医薬品局の承認を受けて1994年に販売が開始された。フレーバーセーバーは,果実が熟するときに働く細胞壁ペクチン(ポリガラクツロン酸)の加水分解酵素であるポリガラクツロナーゼの生成を抑えるために,ポリガラクツロナーゼ遺伝子のアンチセンスcDNAを遺伝子導入して作られた。アンチセンスcDNAは,標的遺伝子のmRNAに対して相補的な配列を持つRNA(アンチセンスRNA)を細胞内で合成させ,翻訳過程を阻害することによって標的遺伝子の発現を抑制するものです。
遺伝子組み換え作物はGenetically Modified Organism(GMO)と呼ばれる。これまでの遺伝子組み換え作物は企業の論理が先行して取り組まれたため,特に食用作物としての遺伝子組み換え作物に対しては社会の拒否反応が強く,遺伝子組み換え作物が社会に受け入れられるためにはPA(パブリック・アクセプタンス Public Acceptance) 「社会的受容性」に対する取組が不可欠である。
現在,遺伝子組み換え作物として日本で受け入れられているものとして1995年に開発された青いカーネーション「ムーンダスト(Moon dust)」を挙げることができる。ムーンダストは1997年から販売が開始され,2005年には全国各地の花店で販売が行われている。2004年に開発された青いバラは次の遺伝子組み換え植物として期待されており,2008年に一般販売が開始される。
 (第34図)
(第34図) 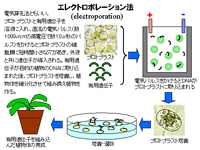 (第35図)
(第35図)
 (第36図)
(第36図) 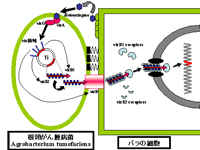 (第37図)
(第37図)  (第38図)
(第38図)
(第39図) (第40図)
(第41図)  (第42図)
(第42図)  (第43図)
(第43図) 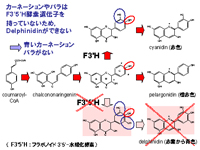 (第44図)
(第44図)
(第45図,第46図,第47図,第48図,第49図)
Ⅶ.知的財産権としての植物育成者権
農業,園芸分野において優良な新品種の作出は,その発展に極めて大きな役割を果たしてきた。新品種の育成には膨大な年月と経費を要し,育成者の労力は多大なものであり,育成された品種は人類の財産として保護される必要がある。品種育成者の権利は,工業特許などと同様に知的財産権の1つとして認知されており,植物育成者権(Plant Breeders’ right (PBR))といわれている。
新品種を育成した育成者の権利を保障し,保護することを目的とした国際的な品種保護条約が1961年にヨーロッパを中心に締結された。1968年には条約加盟国による植物新品種保護国際連盟(UPOV)が発足し,UPOV加盟国は1978年に条約改正を行い,その後1991年3月には加盟20カ国によって再度の改正が行われた。
この改正によって新品種の登録有効期間が延長されたほか,保護対象があらゆる植物の種を対象とされた。我が国では,1978年に農林種苗法から種苗法が改訂され,1982年9月に1978年UPOV改正条約に準拠した改正が行われた。1999年12月には1991年改正条約に準拠してた種苗法として再度改訂された。
ヨーロッパ諸国,南アメリカ,北アメリカ,オーストラリア,ニュージーランド,南アフリカ,イスラエルなどの国では1991年UPOV改正条約に準拠した国内法を制定しており,すべての植物に規制が及んでいる。しかし,中国や韓国をはじめとする先進アジア圏の各国は,1978年の改正前のUPOV条約に準拠した種苗法に基づく法律を施行しており,新種苗法との整合性が取れていない。加えて他の多くの国では1968年当初のUPOV条約にも準拠していない。また,中国や韓国ではバラは指定品目に挙げられてはいるものの必ずしも法律が遵守されておらず,これらの国に新品種の切りバラを輸出した場合,無断で増殖,栽培が行われ,パテント料が支払われず育成者権が保護されない恐れがある。
種苗法は,育種を進める上で関係者が必ず遵守すべき法律であり,種苗法を十分理解する必要がある。