![]()
園芸栽培汎論の講義内容
1.園芸と園芸学の定義
(1)定義と特徴
園芸の歴史は人類が文化を持ち始めたことと同義であり、文化の歴史と同じである。日本における園芸は、「自然をめでる心」や「食糧の確保」と共に発展してきた。花を楽しむ心は自然観を豊かに育むことであり、季節の植物を観賞することは季節感を感じることでもある。すなわち、春のサクラ、夏の早朝のアサガオ、初秋のススキ、ハギ、初冬のサザンカなどは季節感を感じさせられる植物の代表である。また、古来より「春の七草」と呼ばれる「せり、なずな、おぎょう(ハハコグサ)、はこべら、仏の座(タビラコ)、すずな(カブ)、すずしろ(ダイコン)」は春の到来を待ちわびる心と共に、七草粥として食べられる植物でもある。七草粥は季節感を感じると共に、冬に欠乏しがちなビタミン、ミネラルを補給するという効用を期待するものであり、野菜としての園芸の発展とも結びついている。同様に果物にも季節感があり、ウメ、モモ、ブドウ、ナシ、カキ、リンゴなどは季節感あふれる果物といえる。このように、季節と野菜や果物の関係は「旬」と呼ばれる。

現代の園芸技術の発展は、「周年生産」「周年供給」を可能にし、いわゆる「旬」の感覚を希薄にしてきた。しかし、これは野菜や果物がビタミンやミネラルの安定供給という宿命を持ったことから始まることであり、同様に「常に花を愛でたい」という心の癒しも含めて、人間の健全な生活を確保するうえで余儀なくされてきたものである。また、日本人の特性として、「季節感の先取り気質」も大いに影響を及ぼしている。

園芸は大きく分けて、個人が楽しむ「趣味園芸」と、産業として生産農家が行う「生産園芸」に分けられる。「趣味園芸」は植物を育てる楽しみや喜びを味あわせてくれる分野であり、最近ではこの人間としての本質的な行為を利用した医療行為としての園芸療法も注目を浴びている。
これに対して、「生産園芸」は生産者が経済行為として生産した生産物を、市場出荷などを通じて消費者が消費するという産業であり、「趣味園芸」とは一線を画したものとして発展してきている。すなわち、需要と供給の間で行われる園芸であり、「周年生産」「周年供給」などのように必ずしも古来から発展してきた本質的な園芸とは一線を画している部分を持っており、近年では国際間の農産物の流通も極めて盛んとなってきている。
生産園芸の歴史は栽培の効率化の歴史であり,1820年には農業者1人当たり3人の消費者の食料供給を行っていたのに対し,1910年では7人,1982年には78人の消費者の食料供給ができるような生産の効率化がはかられてきた。
「園芸」という用語は英語の「horticulture」の訳語に由来し、「horticulture」はラテン語の「hortus(囲い)」と「cultura(栽培管理)」からなる。漢字の「園芸」の語源は「庭に手を加える」の意味で,ここでいう「庭」とは「庭園」や「果樹園・菜園」なども含む「植物の生育している区域」の意味である。したがって、園芸作物とはこれらの区域に生育する植物で(garden crops),果樹・野菜・花・樹木を含む。園芸作物と対照となる作物は畑作物(主に穀物)や林木である(field crops, forest trees )。
(2)園芸作物
園芸作物は、果樹(fruit trees)、野菜(蔬菜)(vegetable crops)、花卉(flower and ornamental plants)に分けられ、一般に生鮮な状態で利用される。園芸作物(garden crops)は農作物(field crops)と区別され、農作物には食用作物(food crops)、飼料作物(forage crops)、工芸作物(industrial crops)、緑肥作物(green manure crops)が含まれる。
園芸作物の野菜や果実は柔軟多汁で、一般に貯蔵性が低く、カロリーやタンパク質に乏しいが、無機塩類、ビタミン類、食物繊維に富み、人間の栄養上、あるいは嗜好上、日常生活に欠くべからざる食品である。また、花卉は、切花、鉢物などの室内装飾や、庭、公園、街路などの植栽植物として利用され、室内外の環境美化や精神的な安らぎと潤いを与えるものである。
【果樹と野菜】
果樹は,文字通り木本植物(樹木)に結実する果実を指す。しかし,例外もあり,バナナは草本生食物であり,1年生宿根植物であるが果樹に分類される。同様にパインアップルもアナナス科の草本植物ですが,果樹に分類される。
パパイアは草本性の多年草で,木本植物ではないが果樹に分類される。しかし,未熟な状態で収穫された果実は野菜として食用される。
野菜は,食用とする草本植物を指す。しかし,例外としてタラの芽などの山菜は木本植物であるが野菜に分類される。
【果物と野菜】
果物と野菜の区別は極めて難しい問題である。例えば,イチゴは野菜に分類されるが果物であり,メロンも同様に野菜であるが果物である。
果樹と果物は定義が異なる。果樹は栽培学上の分類であり,果物は流通における分類である。メロン農家は,栽培学上の分類では野菜生産農家であり,市場に出荷した段階で出荷されたメロンは果物となる。
(3)園芸学
園芸作物や園芸に関する科学を総称して園芸学(horticultural science)といい、応用科学である。対象作物別に、果樹園芸学(pomology)、野菜(蔬菜)園芸学(vegetable crop science)、花卉園芸学(floricultural science)に分けられる。
園芸学は「園芸」という産業を基盤とした学問であり、産業と一体となって研究が行われる性質を持つ。したがって、産業として成り立たない「園芸学の研究」は本質的に「園芸学」とはいわない。【農学栄えて農業亡ぶ】
園芸学は、これまで対象作物別に体系付けられてきたが、近年園芸作物に共通する理論を基礎とした新たな体系が作られ始め、園芸育種学、園芸繁殖学、種苗生産学、園芸作物生理学、施設園芸学などが体系付けられた。
【園芸学は実学である】
(4)園芸の特徴
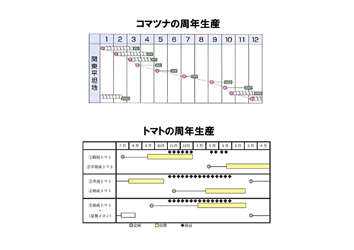
園芸作物は貯蔵性に乏しく、周年新鮮な形で供給される必要がある。周年供給の方法として、(1)年間を通じて作型や品種を変えて周年生産を行う、(2)施設を用いて年中同一環境を維持して周年生産を行う、(3)南北に長い日本の地理的特徴を活用し、季節に応じて適地で生産し、消費地に供給する、(4)貯蔵技術の向上により安定供給する、(5)国際的な流通、などがある。
(1)周年生産を行うために作型や品種を変えて行う必要がある。すなわち夏は日射量が多いが高温であり、冬は日射量が少ない。したがって、冬に栽培を行う場合には低日射量でも生育ができる(光合成効率の高い)品種を選択する必要がある。このように、作型に応じた品種の開発が行われ、例えばスイカでは200品種が存在するといわれている。
(2)施設生産は周年生産を行うためには必要不可欠な栽培技術であり、総面積は約112,000haで、野菜90,000ha、果樹12,000ha、花き10,000haと野菜が最も多く、施設面積は園芸大国といわれるオランダより多い。
(3)地理的特徴を活用した周年供給は、「リレー栽培」とも呼ばれ、北海道と和歌山県でスターチスの切花生産を行い、同じブランド名で首都圏市場に周年供給しています。
(4)貯蔵による周年出荷はリンゴに代表される。11月に収穫されたフジは8ヶ月のCA貯蔵が可能となり、極早生品種の収穫が始まる7月まで出荷される。
(5)国際的流通には、(1)ニュージーランドからのアスパラガス輸入のような南北半球を活用する、(2)冬季の台湾からのタマネギの輸入など日本で栽培不適な気候の時期に輸入を行う、(3)年間を通じて安定した気候の地域(熱帯高地)で周年生産・輸入を行う、などがある。
園芸生産物は貯蔵性が乏しいことから出荷調整が難しく、また農業の性質として気象の影響を受けやすく、需給バランスによる価格変動を受けやすい。また品質による格差が大きく、商品生産としての特徴をもつ。このような特性から、園芸生産では集約的な管理を必要とし、特に施設園芸では最新の技術と設備が導入され、近年では環境制御や潅水施肥などの栽培管理などでのコンピュータの導入が著しい。
バラの国際流通
2.発展の歴史
(1)園芸作物の起源
人間が文化・文明を維持・発展させるためには衣食住の安定が不可欠であり、食の安定的な確保は文化の基盤である。旧石器時代の人類は、野生の動植物を捕獲採取していたが、新石器時代になると野生の植物の中から選抜し、栽培を行うようになり、定住化が行われ始めた。野生植物の栽培化の過程で、有用形質が選抜され、栽培植物化が行われた。
栽培植物の起源については1800年代から多くの研究が行われ、Darwinの「種の起源」(1867)、De Candolle「栽培植物の起源」(1883)を始めとした論文がまとめられた。1900年代に入ってVavilovが「遺伝子中心説」(1928)を唱え、世界8地域を中心とした地域が栽培植物の大きな起源地域であることを明らかにした。
中国 (136種)
ハクサイ、ダイズ、ネギ、ゴボウ、ヤマイモ、ナシ、カキなど
インド、東南アジア(172種)
キュウリ、ナス、サトイモ、ショウガ、トウガン、バナナなど
中央アジア(42種)
ダイコン、ニンジン、タマネギ、ホウレンソウ、リンゴ、ブドウなど
近東(83種)
メロン、レタス、ブドウ、イチジクなど
地中海(84種)
エンドウ、キャベツ、アスパラガス、セルリーなど
アビシニア(エチオピア)(38種)
ササゲ、オクラ、スイカなど
中央アメリカ(49種)
トウモロコシ、インゲンマメ、サツマイモ、シシトウガラシなど
南米(62種)
ジャガイモ、トマト、カボチャ、イチゴ、パインアップルなど
これらの世界的にみた園芸作物の発展と比較して、日本における園芸作物の発展は日本固有の植物の栽培化に加えて、中国などから導入されたものが多い。
(2)日本における園芸の発達
縄文・弥生時代は農作物と園芸作物を区別することはなく、食料の確保としての野生植物の栽培が行われていた。奈良・平安時代になると、ニホンナシ、ニホングリ、フキ、セリ、ウド、ミョウガ、ハス、サクラ、ウメ、ハギ、アジサイ、ノイバラなどの日本自生の植物を栽培化した園芸作物に加えて、中国を経由して渡来してきた作物も多く栽培されていた。カンキツ類、ダイコン、カブ、ニンニク、シソ、ラッキョウ、ナス、キュウリ、ササゲ、ニラ、レアス、エンドウ、キク、ボタンなどが挙げられる。
奈良・平安時代から鎌倉時代になると中国大陸からの貿易が盛んになり、それと共にカンキツ類、ブドウ、アンズ、ウメ、ザクロ、カキなどが伝来した。
鎌倉・室町・桃山時代にはカボチャ、トウガラシ、トウモロコシ、ジャガイモなどの新大陸由来の作物が渡来した。
自給的な生産ではなく、農業としての園芸作物の生産は江戸時代に発達し、都市の発達と共に流通が完備し、商品作物生産が行われるようになった。商品作物生産の発達に従って産地の形成が行われはじめ、大和のカキ、甲斐のブドウ、伏見のモモ、丹波のクリ、紀州のミカンなどの産地が形成された。これまでにはなかった江戸時代の大きな変化として、「伊勢参り」がある。弥次喜多道中にあるように伊勢神宮に地域の代表としてお参りをする「伊勢参り」のもう一つの目的として「品種交換会」があったと言われ、江戸時代後期には地域特有品種が次第に全国に広がり始めるきっかけとなった。
花き園芸では、地域特異的な品種群の形成(嵯峨ギク、美濃ギクなど)やアサガオ、サクラソウ、フクジュソウなどのような特異品種の確立がみられ、世界中でも最も園芸文化が進んだ時代といわれた。
明治時代になると、国策として欧米の園芸の導入が行われ、果樹では米国ブドウ、リンゴ、オウトウ(サクランボ)などが定着した。
第2次世界大戦以前は、多くの野菜品種は固定種であったが、その後F1雑種が多くを占めるようになり、民間育種から企業育種に変化し、種苗会社が台頭し始めた。
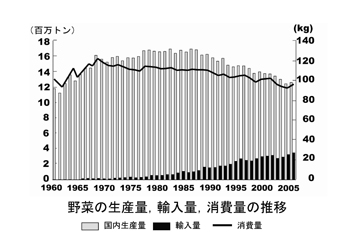
(3)生産と消費
農業総産出額は84,887億円(2005年)のうち、園芸生産は26,762億円(31.5%)で、米の19,650億円(23.1%)、畜産25,548億円(30.1%)と共に3本柱の一つとなっている。園芸作物の生産額は年々増加する傾向にあり、20年間で果樹では2.6倍、野菜では3.5倍、花きでは8.4倍で花き生産の増加が著しい。過去10年間についてみると、野菜の増加は少なく、果樹では減少傾向にあり、花きだけが急激に増加している。
果樹の主要な品目を見ると、収穫量で見ると、温州ミカン、リンゴ、ナシ、ブドウ、カキの順である。これに対して栽培面積では温州ミカン、リンゴに次いでクリ、カキ、ナシ、ブドウの順となっている。
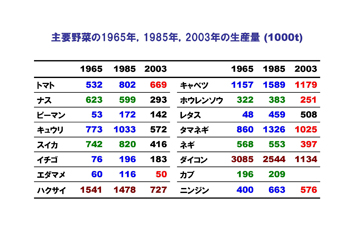
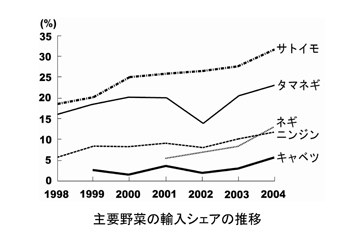
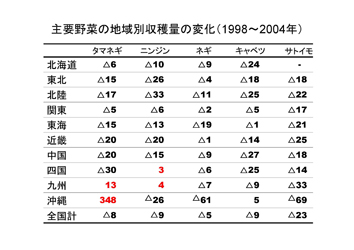
野菜の主要な品目を見ると、収穫量で見ると、ダイコン、キャベツ、タマネギ、キュウリ、トマト、スイカ、ニンジンの順となる。
1965年から1985年の20年間で増加した野菜はトマト、ピーマン、キュウリ、イチゴ、エダマメ、キャベツ、レタス、タマネギ、ニンジンがあり、減少あるいは変動がない野菜はナス、ハクサイ、ネギ、ダイコン、カブなどである。
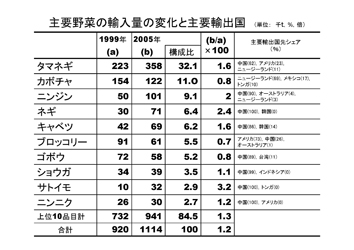
しかし1985年から2003年にかけては、レタスで増加、イチゴが横ばいの他はほとんどの野菜が減少しており、なかでもキュウリ、スイカ、エダマメ、ハクサイ、ダイコンなどは半減している。この要因として、野菜消費量自他員減少に加えて、海外からの輸入野菜の増加が大きな影響を与えている。
花きでは、切花と鉢物に分けられ、切花ではキク、バラ、カーネーション、ユリ、カスミソウの順で生産量が多く、なかでもキク、バラ、カーネーションは世界的にも3大切花品目となっている。
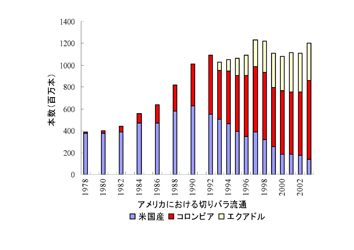
(4)輸出入
2005年の農産物の輸入総額は47,922億円で、輸入総額569,494億円の13.4%であった。輸入国はアメリカ(22.7%)、中国(14.6%)、EU(10.8%)、オーストラリア(7.9%)、カナダ(6.5%)の順である。このうち果実は193万トンで、2,323億円で、アメリカ、フィリピン、ニュージーランド、メキシコ、中国の順で、国内の総出荷量が230万トン(2006)であることから総流通量の46%が輸入されていた。最も多いのはバナナ、グレープフルーツ、キウイフルーツ、オレンジ、レモン、パインアップルなどである。
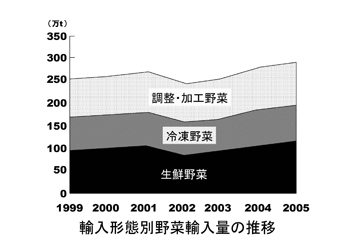
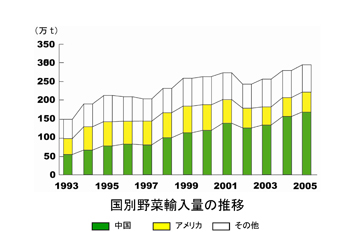
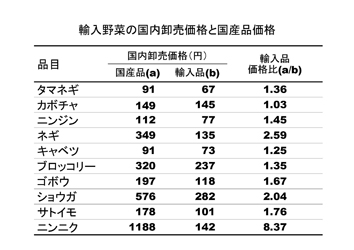
生鮮野菜についてみると、100万トン、955億円で、中国、アメリカ、韓国、ニュージーランド、タイの順で、内訳は、タマネギ、カボチャ、ニンニク、ニンジン、ネギ、キャベツなどである。
冷凍野菜では74万トン、1,020億円で、中国、アメリカ、タイ、台湾、カナダの順で、エダマメ、スイートコーンなどが多い。
生鮮野菜と冷凍野菜の合計は174万トンに達し、国内の総出荷量が1,035万t(2006)であったことから総流通量の14%が輸入されていた。
花き関係では切花が多く、ラン、キク、バラ、シダ、ユリなどが急増している。
3.園芸植物の分類
園芸植物を分類する方法として、自然分類と人為的分類がある。
(1)自然分類
自然分類は植物分類学に基づく分類で、国際植物命名規約によって規定されている。界(Kingdom)、門(Division)、綱(Class)、目(Order)、科(Family)、属(Genus)、種(Species)、変種(Variety)、品種(Form)とする。ここでいう「品種(Form)」とは,分類学上のもので,園芸植物に用いられる「栽培品種」とは異なる。「栽培品種」の英訳として Variery と Cultivarが用いられるが,分類学上の「変種(Variety)」との混同を避けるため,「Cultivar; cv.」を用いる。分類学上の「品種(Form)」は花や葉の斑入りなどの場合に命名される。園芸植物に用いられる「栽培品種(Cultivar)」は農林水産省が監督する種苗法に基づき品種登録されたものをいい,クローン,固定種,交雑種,自然交雑品種などがある。
この中で、属以下の分類は極めて類似性に富み、形態的にももっとも把握しやすい分類である。
クローン(Clonal cultivar):栄養繁殖性の植物で用いられる品種の概念で,野菜ではジャガイモ,イチゴなどがあり,ほとんどの果樹,鑑賞植物ではバラ,球根,セントポーリアなどが有名である。これらの植物は,挿し木,接ぎ木,株分け,ランナーなどで繁殖され,近年では組織培養が用いられる場合も多い。クローンによる変種は,交配後の実生から選抜される場合,遺伝的な突然変異による場合があり,特に後者では「系統(Strains)」あるいは「枝変わり(Sports)」と呼ばれる。例としては,交配によって育成されたリンゴ品種「デリシャス(Delicious)」の枝変わりとして「スターキングデリシャス(Starking Delicious)」,「スタークリムソンデリシャス(Starkrimson Delicious)」,「リチャードデリシャス(Richared Delicious)」などがある。
固定種(Pure-line cultivars):種子繁殖性で自家受粉植物の多くは固定種として品種が作られており,その多くは遺伝的に純系(ホモ)(Homozygous)である。したがって,これらの品種では自家採種が可能である。
交雑種(Hybrid cultivars):種子繁殖性で異なった親系統を交雑することによって作られる品種で,F1雑種品種として有名である。これらのすべては遺伝的にヘテロ(Heterozygous)で,自家採種を行うことが不可能であるため,種苗会社にとっては有利な販売を行うことができることから,ほとんどの野菜あるいは種子繁殖性の花の品種はF1品種となっている。
自然交雑品種(Open-pollinated cultivars):キャベツやホウレンソウなどのように自殖弱勢が強い植物では,遺伝的に純系の個体を作出すると採種能力が著しくて以下するため,ある程度の範囲で形質が異なっている親同士を自然交雑して品種を育成する。
植物の名前には、園芸名、和名、学名がある。例えば園芸名サルビアの和名は、ヒゴロモソウで、この名前はSalvia属に属する植物の中で園芸的に利用されているものの総称で、Salvia splendens F.Sello,S. farinacea Benth. ,S. horminum L.などが含まれ、これらの中に品種名(栽培品種名)として、セント・ジョンズ・ファイア-、ホット・ジャズなどがある。
一般に園芸植物の命名には二命法が用いられており、属名と種名が併記される。園芸植物の中でも花きは種間雑種や属間雑種が多く作られているため、その命名法について述べる。
種間雑種 Camellia japonica とCamellia saluenensisを交配した品種'Donation'は、Camellia japonica X C. saluenensis 'Donation' と表記する。ただし、この交配種に新たな種名 williamsii が与えられている場合には、CamelliaXwilliamsii 'Donation' と表記しても良い。
属間雑種 Amaryllis belladonnaとCrinum moorei の交配種は、Amaryllis belladonnaXCrinum moorei 'Elliy' と表記されるが、この交配属名としてアマルクリナムが与えられているばあいには XAmarcrinum 'Elliy'とする。このように属間雑種には両親の属名の一部が組み合わされる場合が多い。
細胞融合による種間雑種あるいは属間雑種の場合には、通常の雑種で用いられるXではなく(X)が用いられる。Nicotiana glauca (X) Nicotiana langsdorffii やSolanum tuberosum (X)Lycoperusocon esculentum(ポマト)など。
果樹のなかで重要な科はバラ科で、リンゴ、モモ、ウメ、オウトウ、ビワなどがバラ科に属する。
野菜では、アブラナ科、マメ科、ナス科、ウリ科、キク科、セリ科、ユリ科に属するものが多く、主要7科と呼ばれる。
(2)人為的分類
食用植物(Edibles)
野菜(Vegetables)
果菜(Fruit vegetables) トマト,キュウリ etc.
花菜(Flower vegetables) カリフラワー,ナバナ etc.
葉菜(Leaf vegetables) レタス,キャベツ,タマネギ etc.
茎菜(Stem vegetables) アスパラガス,コールラビー etc.
根菜(Root vegetables) ダイコン,ニンジン,サツマイモ etc.
果樹(Fruits)
落葉果樹(Deciduous)
核果類(Stone fruits) モモ,オウトウ,ウメ
仁果類(Pome fruits) リンゴ,ナシ
堅果(Nuts) クリ,クルミ,アーモンド
小果類(Small fruits)
液果(Berries) イチゴ,ブルーベリー
木イチゴ類(Brambles) ラズベリー,ブラックベリー
つる性(Vines) ブドウ,キウイー
その他 カキ,イチジク,ザクロ
常緑果樹
草本性果樹(Herbaceous fruits)
バナナ,パパイヤ,パイナップル
木本性果樹(Tree fruits)
カンキツ類(Citrus) オレンジ,ミカン,グレープフルーツ
その他 アボカド,マンゴ,マンゴスチン
鑑賞植物(Ornamentals)
花,観葉植物(Flowers and Foliage plants)
一・二年生草(Annuals, Biennials)
マリーゴールド,ペチュニア
多年草(Perennials) キク,シャクヤク
球根(Bulbs, Tubers)
鱗茎 チューリップ,スイセン、ヒアシンス(有皮鱗茎)
ユリ(無皮鱗茎)
塊茎 アネモネ、ベゴニア、シクラメン
球茎 クロッカス、フリージア、グラジオラス
根茎 カンナ、アイリス、シャクヤク、ジンジャー
塊根 ダリア
花木(Ornamental trees) バラ,サルスベリ,キンモクセイ
観葉植物(Foliage plants) ポトス,ドラセナ,シダ類
ラン類(Orchids) カトレア,デドロビウム
サボテン・多肉食物(Cacti, Scculent plants)
サボテン,金のなる木
景観植物(landscape)
芝(Lawn, Turf)
グランドカバー,ツル性植物(Ground covers, Vines)
庭木
4.形態
(1)栄養器官の形態
植物体=シュート(shoot)+根(root)
シュート=葉(leaf)+茎(stem)
茎=節(node)+節間(internode)
根=定根(morphological root)+不定根(adventitous root)
定根=主根+側根
(a)茎
短縮茎(dwarf stem)、直立茎(erect stem)、巻きつき茎(twining stem)、匍匐茎(creeping stem)
木質の発達した木本と発達しない草本にわけられる。
草本は1年生植物(annuals)、2年生植物(biennials)、多年生植物(perennials)
内部構造は、表皮、皮層、内皮、内鞘、中心柱
(b)葉
葉の変態
?葉球形成
?鱗茎形成
(c)根
初生根、主根、種子根
定根と不定根、側根
根毛とその寿命
根の変態
?貯蔵根
(2)生殖器官の形態
(a)花と果実
子房の着生位置と果実の構造(カキ、ミカン、モモ、リンゴ、メロン、イチゴ)
雌雄異花(雄花と雌花)、両性花、雌雄同株、雌雄異株
花器と果実
(b)種子
受精と種子形成
胚乳種子と無胚乳種子
5.成長と発育
(a)植物の生活環
植物のage(幼若相と成熟相)
(b)種子の発芽と休眠
種子の発芽には「水、温度、酸素」が必要である(ただし、イネ科の水生植物のなかには酸素を必要とせず、無気呼吸によって発芽するものもある)。この発芽3要素が欠如するなどの外的環境条件によって発芽が抑制されている状態を「多発休眠(強制休眠)」という。これとは異なり、内生発芽抑制物質による発芽阻害や吸水阻害などのによって発芽できない状態を自発的休眠という。
発芽に関わる外的要因として光を必要とするものがあり、発芽に光が必要なものを「光発芽性種子(好光性種子)」という。この仲間には野菜ではレタス、ミツバ、ゴボウ、シュンギクなどのキク科植物があり、花きではAdonis, Alisma, Bellis, Capparis, Colchicum, Erodium, Fagus, Genista, Helianthemum, Iris, Juncus, Lactuca, Magnolia, Nasturtium, Oenothera, Panicum, Resedea, Salvia, Suaeda, Tamarix, Taraxacum, Veronicaなどがある。逆に暗黒化で発芽しやすいものとして野菜ではスイカ、カボチャなどのウリ科植物やユリ科植物があり、花きではAilanthus, Aloe, Cistus, Delphinium, Ephedra, Evonymus, Forsythia, Gladiolus, Hedera, Linnaea, Mirabilis, Nigella, Phacelia, Ranunculus, Silene, Tamus, Tulipa, Yuccaなどがある。光感受性種子は、吸水後初めてフィトクロームを介して光に反応するようになる。光反応性種子が必要とする光照射時間は極めて短く、レタスで1~2分といわれ、好光性種子の発芽は660nm付近の赤色域の光で促進され、730nm付近の遠赤色域の光で抑制される。赤色光と遠赤色光を交互に照射すると、最後に照射した光の波長によって発芽の誘導の有無が決定される。(p106の表22)
成熟した種子の生理活性状態はほとんど休眠状態にある。休眠は外的要因に起因する休眠(強制休眠または他発休眠)と内生的な要因(生理的な要因)や種子の構造的な要因によって発芽ができない自発休眠に分けられる。種子の休眠は、春と秋の気候条件は似ているため、秋前に成熟する種子が秋の気候に反応して発芽するのを避けるための予防策であり、また翌年一斉に発芽した場合に気象異常で全滅するのを防ぐため数年間発芽できないようなシステムでもある。
休眠には「生理的休眠(自発休眠)」と「物理的休眠(多発休眠)」がある。
自発休眠には、胚の活性低下と胚以外の種皮などの構造に基づくものとに分けられる。胚の活性低下に起因するものとして、?胚の未成熟、?胚の代謝能の阻害などがあり、種皮などの構造によるものとして、種皮の水不透過性、種皮の酸素や二酸化炭素不透過性、種皮の機械的抵抗がある。
?胚の未成熟:アメリカヒイラギやイチョウ、ラン類では種子が成熟したときには胚がまだ未成熟で、追熟過程が必要。【休眠打破処理】20~30?で維持し、追熟させるか、あるいは無菌播種を行い人工的に培地から養分を供給する。
?発芽阻害物質:果実や種皮に発芽阻害物質が含まれている。トマトやホウレンソウの果実には阻害物質が含まれており、種子を水洗すると発芽可能となる。【休眠打破処理】1晩程度、流水で水洗する。果皮をよく洗って取り除く。
?植物ホルモン(アブシジン酸:ABA):種皮に含まれ、種子が発芽するとき必要なα-アミラーゼの活性を抑える。ジベレリンとは拮抗作用を持つ。【休眠打破処理】水洗することで除去ができる。ジベレリン処理を行う。
?発芽促進物質の活性低下:種子中の発芽促進物質の活性が低く、発芽ができない。低温・高湿処理(層積処理)によって活性が高まる。発芽促進物質は幼根のなかにあり、低温・高湿処理によって不活性型から活性型に変化する。【休眠打破処理】低温・高湿処理:高湿度条件で、0~10?(5?がベスト)に維持する。湿ったバーミキュライトやピートモスに種子を入れて、冷蔵庫で保存する。通常は3ヶ月程度。(Viburnum(ガマズミの仲間)では果実の成熟段階では幼根が未成熟なため、20~30?条件で幼根が発達し、その後低温・高湿処理を行う。)
種皮などの構造による休眠は「硬実」とも呼ばれ、種皮が硬かったり、厚かったり、あるいは脂質やタンニンが多量に含まれたりして、吸水阻害が起きる。自然界では、土壌中の微生物が種皮を徐々に分解したり、風化することで次第に種皮が薄くなり、吸水可能となる。【休眠打破処理】ヤスリなどで種皮を削ったり、濃硫酸で種皮を溶かす。Gleditsiia(サイカチ)などでは熱湯や熱アルコ-ルで処理することで、脂質やタンニンを除去し、吸水を促す。
(c)開花
栄養生長を行っている茎頂生長点が花芽分化し、花芽の発育が完了すると(3年後期の園芸生産工学で解説)、開花が始まる。開花の引き金となる外的要因として、温度、日長、光強度が挙げられる。日長に関しては、開花前日の日没後の暗期継続時間が関係し、アサガオでは約10時間、カボチャでは6~7時間、スイカでは約8時間で開花する。チューリップ、クロッカス、マツバボタン、カタバミなどでは温度の変化が関係し、現在より高温になると開花し、低温になると閉花する。ツキミソウ、マツヨイグサなどでは光強度が関与し、開花時刻は日没時刻とほぼ一致する。
(d)受粉・受精
開花した子房内には胚珠があり、胚珠には胚のう(卵細胞、極核、助細胞、反則細胞)がある。受粉した花粉は柱頭で発芽し、花柱内部を花粉管が伸長する。発芽前の花粉には大きな花粉管核と小さな生殖核の2核が存在するが、花粉管が伸長すると生殖核は花粉管内で2個の精核に分裂し、花粉管の先端に花粉管核が位置し、その後方に2個の精核が位置する。花粉管は珠孔を通じて胚のう内に進入し、花粉管膜が破れて2個の精核が放出され、卵細胞(n)と極核(2n)と受精して卵細胞は胚を形成し、極核は胚乳となる。
正常な生殖器官を持っていても自家受粉した場合には受精が行われない場合があり、これを自家不和合性という。バラ科の果樹のリンゴ、ナシ、オウトウ、スモモ、アーモンド、ウメやアブラナ科野菜でみられ、これらの作物では異なる品種の花粉を交配する必要がある。これには複対立遺伝子が関与しており、S遺伝子の形質が同一であるか否かが自己・非自己の認識と関係している。
一般に果実の肥大は受精によって種子発育が開始することで行われるが、なかには種子が形成されなくても果実が肥大する現象が認められ、単為結果という。単為結果には2つのタイプがあり、1つは受粉や外的刺激がなくても果実が発育する自動的単為結果(カキ、カンキツの一部、イチジク、キュウリなど)で、もう1つは受粉や外的刺激によって単為結果する他動的単為結果である。他動的単為結果には、リンゴで見られる霜などの物理的刺激やトマトのオーキシン、ブドウのジベレリンのようなホルモン剤などの化学薬剤の刺激がある。単為結果しやすい果実では、花蕾から幼果にかけての子房内のオーキシンなどの植物ホルモン活性が極めて高いことが知られており、本来、胚や胚乳の発育に伴って生合成される植物ホルモンが果肉組織内で生合成されることで種子が未形成でも果実が発育する。
(e)果実の発育
果実の発育曲線は、単一S字曲線を示すリンゴ、ナシ、カンキツ、バナナ、トマト、ナス、メロン、スイカと二重S字曲線を示すモモ、オウトウ、ウメ、アンズ、イチジク、カキ、ブドウに分けられる。
果実の発育は、初期の細胞分裂による果肉細胞数が増加する時期(細胞分裂期)と個々の果肉細胞が肥大したり細胞間隙が増大する時期(細胞肥大期)に分けられる。ナス科果菜類、ウリ科果菜類、イチゴなどでは主要な細胞分裂は開花前に終了し、果実の細胞数は開花期にはほぼ決定され、開花後は主として細胞肥大によって果実の肥大が行われる。
6.園芸植物の育種
園芸植物の育種には導入育種,分離育種,交雑育種,倍数性育種,突然変異育種,細胞工学育種,遺伝子組み換えがある。
(1)導入育種
他地域から品種を導入し,栽培地域に適した品種を選抜する。大陸東岸と西岸では気候が異なり,大陸西岸の花き冷涼乾燥気候のヨーロッパで育種された品種は,大陸東岸の高温多湿な日本や中国では病気にかかりやすく,適応性が低い品種が多い。ヨーロッパで育種された品種は日本に一端導入して適応性の検定を行い栽培する必要がある。
(2)分離育種
特定の形質に注目して集団から分けて採種を続け、固定度を高めていく育種方法で,特に自家不和合性を持っている作物で行われる。この方法で育種した品種は固定種と言われる品種となる。
(3)交雑育種
異なった形質を持った複数の個体間で交配をし、その後代から望みの特徴を合わせ持った個体を選抜して固定する。
雑種強勢育種 固定された系統同士を組み合わせて、両者の能力を足した以上の能力を発揮させる。
倍数性育種 倍数体の巨大性、不稔性などを利用する。
突然変異育種 物理化学的な手段によって、時には組織培養による変異体出現を促進する手段によって、積極的に新しい変異を作り出す(培養育種を含める)。
また、最近になって盛んになってきた、染色体そのものを操作して遺伝特性を変化させようとする細胞工学育種や、DNAレベルで特定の遺伝子のみを除去したり、結合させたり、組換えたりする形質導入育種まで、育種手法は発展しました。
7.種子繁殖
採種した種子は発芽促進と発芽勢向上を目的として、化学的、物理的、生物的処理(種子処理)を行う。
1)種子発芽の生理
種子は成熟につれて含水率が10%程度まで減少し、この脱水過程で代謝活性は急激に低下し、貯蔵性を獲得する。乾燥した種子は吸水によってタンパク合成ややATP生成を開始するが、この吸水過程は3段階に類別できる。第1吸水過程は種子の生死に関わらず行われ、いわゆる物理的吸水過程である。第2吸水過程では、含水率はほとんど変化しないか、わずかに増加し、この間に生理的な種々の代謝変化が見られ、生理的吸水過程である。第3吸水過程は、幼根の発生に伴う吸水過程で、成長的吸水過程ともいい、この吸水過程で植物体は急速に成長する。
種子の発芽には「水、温度、酸素」が必要である(ただし、イネ科の水生植物のなかには酸素を必要とせず、無気呼吸によって発芽するものもある)。この発芽3要素が欠如するなどの外的環境条件によって発芽が抑制されている状態を「多発休眠(強制休眠)」という。これとは異なり、内生発芽抑制物質による発芽阻害や吸水阻害などのによって発芽できない状態を自発的休眠という。
発芽に関わる外的要因として光を必要とするものがあり、発芽に光が必要なものを「光発芽性種子(好光性種子)」という。この仲間にはレタス、ミツバ、ゴボウ、シュンギクなどのキク科植物があり、逆に暗黒化で発芽しやすいものとしてスイカ、カボチャなどのウリ科植物がある。
2)種子処理
園芸生産において、発芽に関わる形質として「発芽率」と「発芽勢」がある。発芽率は播種した種子の発芽した割合で示す。発芽勢は発芽の斉一性を示す指標で、「T90-T10」で示す(T90:最終発芽率が90%に達した日数、T10:発芽率が10%に達した日数)。
発芽率、発芽勢を良くするために種々の処理を種子に行う。
(1)病害防除を目的とした薬剤処理
種子伝染性病害を防除する目的でチウラム剤やベノミル剤を種子に粉衣する。購入した種子がこれらの薬剤で赤色や緑色を呈する。トマトではウィルス防除を目的として第3リン酸ナトリウム処理を行う。
(2)病害防除を目的とした乾熱処理
メロン、スイカなどのウリ科植物ではキュウリ緑斑モザイクウィルスの防除を目的として70~73℃で3~4日処理を行う。
(3)発芽促進のための低温処理
自発休眠を行う植物では、採種後一定期間自発休眠が見られ、これを打破するために5~10℃の湿潤条件で低温処理を行う。この間に成長抑制物質であるアブシジン酸が減少し、成長促進物質のジベレリンやサイトカイニンの増加が見られる。
(4)発芽促進のための薬剤処理
チオ尿素や硝酸カリウムを処理する。これらは呼吸反応の解糖系の活性化に関与しており、第2吸水段階での呼吸活性の増加を促し、発芽勢を高める。
(5)発芽促進のための剥皮処理
種皮や果皮が厚く、これらが第1吸水過程の物理的吸水を阻害している場合に、種皮や果皮を物理的に除去したり、薄くする処理で、ホウレンソウやシュンギクでは種皮(果皮)を完全に除去し、「ネーキッド種子」として市販されている。また、種皮(果皮)に物理的に傷を付ける処理(磨傷処理)を行う場合もある。
(6)発芽の斉一性を高めるための高浸透圧処理
プライミング処理(オスモプライミング処理)といい、吸水を行わせて第2吸水段階の代謝反応を行わせるものの、高浸透圧剤でそれ以降の代謝反応を抑制させる処理で、この処理を行った種子は第1吸水過程(物理的吸水)を行った後、速やかに第2吸水過程を経て発芽に至る。このため、発芽勢が著しく高まる。
(7)機械播種のための被覆処理
植物の種子の形態は千差万別で、大きさも極めて小さいものがある。プラグ苗生産においては、播種は自動播種機を用いて行うため、機械播種に適するように粘土物質や高分子化合物を被覆して形状を球状やラグビーボール状に整形したり、極小の種子を大きくする処理を行う。これらの種子は「コーティング種子」として市販されている。また、発芽促進や発芽後の成長促進を目的として、これらの被覆資材に殺菌剤や肥料などを混合したものもある。
(8)播種の省力化のための処理
水溶性、あるいは生分解性シートをテープ状にしたものの間に種子を挟み、ひも状あるいはテープ状にした「シードテープ」として市販されており、大面積の圃場においてトラクターなどでこのテープを畑に設置し、その後潅水をするなどの機械化技術として用いられる。北海道でのタマネギ栽培において活用されている。
(9)高分子化合物による流体播種処理
高分子ゲル化剤に種子を混入・拡散させ、一定量ずつゲル剤を滴下することで播種を行う「流体播種」を行う。種子は吸水させ第3吸水過程(幼根の発芽後の種子)のものを混入させることで、播種後の定着が極めて速く行われる。
3)セル成型苗生産(プラグ苗生産)
セル成型苗とは逆円錐形や逆四角錐状の鉢(ポット)をたくさん連結してトレイ状にしたトレイ(セルトレイ)で育苗した苗のこと。トレイの素材はポリスチレンや発泡スチロールが多い。その鉢一つ一つを「セル」と呼ぶ。セルの底には水抜き穴があり,直接地面や水の溜まるような場所から隔離することによって,その穴からは根がでない様にする(エアープルーニングという)。このことによって苗は限られた一定量の培養土しか得られず,根もセルの中でしかのびないため,均一な苗ができ,根が培養土の周りをぐるぐると回り(根鉢の形成),セルから苗が抜けるようになる。培養土は抜けやすくするためや,水分を多く含ませるためなどの理由でピートモスやバーミキュライトといった軽い素材が使われる。プラグ苗は1970年代にヨーロッパで開発され、アメリカで大きく発展し、世界に広くひろがった。プラグ苗の由来は、プラグトレーにあけられた孔(セルともいう)が楔型をしており、ここで育てられた苗はしっかりした根鉢が形成されているため、セルから引き抜いても根鉢が崩れず、定植する際に植え穴に楔(プラグ)を打ち込むように差し込む(電気のコンセントにプラグを差し込むように)ことで定植操作が完了することから付けられた。野菜など大規模圃場にプラグ苗を定植する場合には、プラグ苗定植機を用いて圃場に定植される。プラグ苗定植機は専用のプラグ苗を用い、作業能率は10a当たり2~3時間程度である。レタスなど全面マルチ栽培を対象にした場合には、ポリマルチにバーナやヒータで穴をあけ、プラグ苗を自動給苗して植え付ける。
プラグ苗の長所は、①大量に均一な苗ができる。
②苗が軽い。③定植動機械化がしやすい。④手植えの場合プラグを差すように植えるので植えやすい。
などがある。その反面、欠点として、①培養土が少ないので,育苗中や移植後に乾燥しやすく,かん水が多く必要で,かん水が多すぎると徒長することがある。②移植適期が短い。一定の時期が過ぎると根が老化し,活着が悪くなる。③根鉢ができない品目(玉ネギなど)はトレイから苗が抜き取りにくい。④根鉢ができると植えてからも影響のでる品目(ハクサイなど)は移植後の根張りが悪くなる。⑤培養土がピートモス主体のため,極端に乾燥すると水をはじいてしまう。炎天下の日が続く時期に移植したと場合,浅植えで培養土から水分が蒸発すると,潅水しても吸水しないで枯死する場合がある。などが挙げられる。
(1) プラグ苗生産システム
プラグ苗生産の発達に伴って栽培と育苗の分業化が進み、プラグ苗生産専門の企業が現れた。農家(栽培者)はプラグ苗生産会社から苗を購入し、プラグ苗生産会社は年間を通じて苗生産を行っている。プラグ苗生産会社では、播種は「全自動播種機」を用いて行われている。また、プラグトレーへの培土の充填も自動化されており、プラグトレーへの培養土の充填→播種→覆土→潅水、などの操作がベルトコンベアーの上で自動的に行われ、その後、恒温・恒湿で照明装置のついた「発芽室」で数日間の発芽処理を行った後、完全自動環境制御された育苗温室内で育苗される。
栽培農家は、定植時期を指定してプラグ苗生産会社にプラグ苗の注文を行い、プラグ苗生産会社は出荷期日から計算して、播種を行う。すなわち、播種から育苗までの過程は全て管理された環境下で行うことでプラグ苗の生育をコントロールして苗の出荷時期を計算する。プラグ苗の出荷は宅配便を用いて行われ、プラグ苗は栽培農家まで直接配送される。
一般に園芸作物の定植期は、初春と初秋に集中することが多く、プラグ苗生産会社では生産の効率化を図る目的でプラグ苗の貯蔵を行う。貯蔵は、照明下の低温で実施され、1~3ヶ月に及ぶ場合もある。
トマトなどの果菜類では、プラグ苗を用いて接木を行って生産性を高める方法も開発されている。
8.栄養繁殖
園植物の栄養繁殖法は、①挿し木、②接木、③株分け・分球、④組織培養、がある。
1)挿し木
(1)挿し木繁殖の意義
植物の根は、発芽の際に幼根由来の初生根とそれ以外の根(初生根や2次根、茎などの器官から発生する根(不定根):adventitous
root)からなり、挿し木は初生根以外の器官から不定根を発生させて、植物体の再生を図る方法である。不定根を発生させる器官として、茎、葉、根を用い、その違いによって、「茎挿し」「葉挿し」「根挿し」などがある。植物は分化全能性を持っているため、全ての器官から発根する能力を持ち備えている。
枝の先端で発生した突然変異個体などを増殖する方法として「茎挿し」が一般に行われ、サツキの色変わり変異個体などのように自根苗の増殖が有名である。この他に果樹では耐病性、耐寒性の付与を目的とした台木の増殖なども行われる。この場合には次項の接ぎ木繁殖を併用する。
◎挿し木の短所
(1) 作物や品種によっては挿し木の難しいものがあり、また、親木の年齢が高まるにつれて発根能力が著しく低下する。
(2) 挿し穂を大量に準備することが難しい。
(3) 穂木の状態や挿し穂の条件によって活着率が変化する。
(4) 作物や品種によって側枝性が残り、心立ちが悪く、樹形が乱れる場合がある。
(5) 高齢木の挿し穂の場合や品種によっては初期生育が遅れる場合がある。
(6) 根が側根性となり、発根方向にムラができる場合がある。
(7) 開花が早くなる場合がある。
(2)挿し木の生理
(1) カルス形成と発根
挿し穂の切断面において、傷を受けた細胞は癒傷ホルモンともいわれるオーキシンを分泌する。また、茎の上部から重力移動で下に転流される生長ホルモンのオーキシンがさらに加わって癒傷組織ともいわれるカルスを形成する。カルスは形成層や師部から発達し、これによって挿し穂の切り口は病菌からある程度保護されるとともに、切口からの有用物質の流失も防がれる。形成されたカルス内に通導組織(木部維管束)が分化し、茎の木部維管束と連絡すると同時に根原基(根の基)の分化が始まる。しかし、厳密にはカルス形成と根原基の形成とは無関係であり、カルスは切口の保護のためには必要であるが、発根過程においては必ずしも不可欠とはいえず、過度のカルス形成は発根を阻害することが多い。
根原基の分化は、形成層の細胞分裂に伴って形成され、基本的にはカルス形成とは無関係な現象といえるが、挿し木当初はカルス形成と同調して根原基が分化し、切断部位からの発根がみられるが、その後は茎から直接根原基が分化し始める。根原基の分化は生長ホルモンのオーキシンによって促進され、発根促進剤として使われるオキシベロンはオーキシンの一種のインドール酪酸の製剤であり、ルートンはナフタレン酢酸アミドである。
◎オーキシン
ダーウィン(進化論の著者)親子によってオーキシンの存在がとなえられ、1913年にデンマークの科学者ボイセンーイェンセン(Boysen-Jensen)やオランダのベント(Went)によって屈光性が、オーキシンの特異的作用に基づいていることが明らかにされた。現在、オーキシンにはインドール酢酸(IAA)、インドール酪酸(IBA)、ナフタレン酢酸(NAA)、2,4-ジクロロフェノキシ酢酸(2,4-D)などが用いられている。
(a).オーキシンの生理作用
オーキシンの植物体内移動は、重力に対する正の極性移動と蒸散による樹液の移動と共に行われる負の移動である。正の極性移動では樹皮の皮層部を、負の移動は木部を通ることが知られている。生理反応には発根、カルスの分化、屈性、伸長伸長、維管束分化、果実の肥大生長促進、頂芽優勢、エチレン合成促進(落葉促進、アナナス科植物での花芽分化促進)などがある。これらの作用はオーキシンの物質の種類によって強弱が異なり、インドール酪酸(IBA)は不定根形成が高く、ナフタレン酢酸(NAA)は次いで不定根形成能力が高い。同様にカルス形成能力は2,4-ジクロロフェノキシ酢酸(2,4-D)が著しく高く、ナフタレン酢酸(NAA)は次いで高い。
(b)IAAの合成、分解
色々なオーキシンの中で天然物質として存在するオーキシンはインドール酢酸(IAA)で、IAAはトリプトファンから生合成され、IAA酸化酵素(パーオキシダーゼの一種)によってIAAは速やかに分解される。また、IAAは熱分解や光分解しやすい性質を持っている。
(3)植物の 発根能力
植物の発根能力は、植物体の齢(若さ)と密接な関係があり、齢が若いほど発根能力が高い。この植物体の齢は幼若性とも呼ばれ、若い齢ほど幼若性が高いと表現する。
◎幼若性
幼若性はjuvenilityともいい、実生のものが最も幼若性が高く、樹齢を経るに従い幼若性が低くなる。幼若期の植物の特徴は、旺盛な栄養生長を行い、花芽分化などの生殖生長が抑えられる。果樹などの樹を対象とした場合には、先端の枝ほど花芽の形成が良く、栄養生長の度合いが低い。これに対して幹に近い部位から発生した枝では栄養生長が旺盛で、いわゆる徒長枝と呼ばれる新梢が発生し、その徒長枝では花芽分化が抑えられる。従って、樹全体の幼若性を分類すると、幹(根)に近い部分ほど幼若性が高く、先端に近いほど幼若性が低くなる。
これらのことから、穂木の採取にあたっては、通常に生長した先端の枝を穂木として用いるより親木の幹から発生した枝や剪定後に発生した枝を穂木とした方が発根率が高い。また、親木の年齢と発根能力との間にも密接な関係があり、栽培年数を経るに従って発根率は低下する。従って親木は栽培年数の短いもの程良く、何年も栽培したものでは発根率が低くなる。同様に、花芽の着生した枝を穂木とすると、発根率が著しく低下する。
◎植物の自発休眠
植物、特に温帯性木本植物は自発休眠を行う。一般に8月から自発休眠が始まり、12月には自発休眠が終了する。自発休眠は植物ホルモンのアブシジン酸(ABA)が密接に関与している。7月下旬から8月下旬にかけて展葉している葉でアブシジン酸が生合成され、基部にある芽の鱗片に多量に蓄積される。この鱗片に蓄積されたアブシジン酸の作用によって側芽の生長点の成長が強く抑制され、気温や湿度などの外的条件が良好であっても萌芽することができない。このように植物体内の抑制物質によって萌芽できない状態を自発休眠という。これに対して12月から2月にかけての低温期のように、成長に外的環境が適していないために萌芽できない状態を多発休眠という。
モモ、リンゴなどの落葉果樹では、5月から7月にかけては発根能力が高く、容易に挿し木が可能であるが、8月になるとアブシジン酸の影響を受けて発根率が著しく低下し、12月まで挿し木が困難となる。12月以降については自発休眠が解除されるため、再び発根率が上昇する。
①穂木の採取前に、親木を剪定(切り戻しやピンチ)し、そこから発生した徒長枝や切り戻し枝を用いる
②親木は何年も栽培せず、更新する
③花芽が分化した枝を穂木に用いない
④自発休眠期の材料を用いない
(4)挿し木の一般的留意点
挿し穂は、根圧の強い力による水分の補給がないため、乾燥には著しく弱い。また、切り口は表皮で覆われておらず、内部組織が露出した状態であるため、病菌が進入しやすく、抵抗性も著しく低い。さらに、新たな養分(肥料成分に加えて光合成など)の補給がなく、挿し木と同時に発根、萌芽などの生存維持のための消耗がはじまるが、これらの生長のための養分は穂木に蓄えられている養分のみに依存している。特に、発根は新たな組織を形成するため、多大なエネルギーに加えて、新たな細胞を形成するための養分も必要となる。従って、挿し穂の中に含まれている養分の多少は挿し木の発根率に大きく影響を及ぼす。また、挿し床は切口の水分管理や病害の管理などの面から重要な要素である。
発根が困難である場合の理由として、①乾燥に弱い、②挿し穂が腐りやすい、③生長ホルモン(オーキシン)の体内活性が低い、④発根阻害物質が体内に含まれる、⑤切り口から酸化しやすい(褐変しやすい)、などがある。従って、挿し木にあたってはこれらの悪条件を極力少なくする必要がある。
●乾燥の防止
(1)挿し穂の採取から挿し木まで
挿し穂を親木から採取し、挿し木を行うまでの過程における乾燥の程度と枯死との関係をみると、穂木の水分率が20%程度の減少であった場合にはほとんど枯死はみられない。また、水分減少率が30~40%(作物によっては50%程度まで)であった場合でも枯れることはなく、その後に水が補給されれば発根能力が大きく低下することはない。
挿し穂の採取後の管理方法として、乾燥を防ぐ目的で水に浸漬することが奨励されている。しかし、軟弱な若葉を付けており蒸散が著しく高い場合や水揚げの特に悪い樹種、発根を阻害する有害物質を含んでいる場合、あるいは酸化酵素の活性が高く褐変が著しい場合には、穂木を採取後速やかに水に浸漬する必要があるが、それ以外では水に浸漬する処理は病害の蔓延を助長し、カルスの形成を阻害する場合があることから、水に浸漬しない方が良好な結果が得られる。また、水の浸漬は挿し穂内のオーキシンを流出させる。
◎挿し穂の管理方法
①直射日光には当てない。 日向と日陰の水分蒸散量は大きく異なり、日向の蒸散量は日陰の4~20倍となる。
②高温条件で放置しない 穂木の呼吸を促進し、穂木に含まれるエネルギーの損失を招き、発根が阻害される。
③水に浸漬しない 挿し穂の水への浸漬は、疫病やピシウム病などの病害の蔓延を助長し、オーキシンの流出により、カルスや不定根形成を阻害する。
④風を当てない 過度の風は過剰な蒸散を盛んにするため、風を当てない。
⑤湿度を保つ 蒸散を抑制するため、湿度を高くする。しかしビニルなどで密閉してはいけない。
⑥低温に保つ ある程度穂木を保存する場合には、葉での呼吸を抑え、湿度を高めるために、低温を保ち、できれば湿らせた紙シートと穂木を交互に層状に重ねる。
(2)挿し木後の乾燥防止
挿し穂の吸水は、主として切口から行われ、茎表面からもわずかながら吸収される。これに対して、葉からの蒸散は普通に行われるため、挿し木当初は蒸散を制限する処理を行う必要があり、挿し木における乾燥・枯死は、茎の切口からの吸水と蒸散とのアンバランスによって生じ、挿し木後の乾燥は、著しく発根率を低下させる。
①通常は挿し穂の葉面積が大きい場合には、葉を切除する必要がある。
②寒冷紗で遮光を行い、遮光率は季節によって変える。(強過ぎる遮光は、光合成を阻害し、発根後の生育を阻害する)
③寒冷紗での遮光によって温度上昇が妨げられる。できればパット&ファンなどの装置による冷房を併用する。
④ミスト装置や密閉などで湿度を高める。
⑤過度の送風を行わない。
●挿し穂のステージの選択
挿し穂は、母木から切り離されて養分や水分の補給がなくなったにもかかわらず、挿し付け当初は下からの充分な養分や水分の補給があることを前提としたような盛んな蒸散や勢いのよい新芽の生長を行おうとする。従って、挿し穂の芽の活性が高い部位を用いると、発根がみられないにも関わらず盛んな芽の伸長がはじまり、一見挿し木が成功したかにも見えるが、実際には好ましくない現象といえる。挿し穂の芽の動き始めたものは、芽の生長に多大なエネルギーを必要とするため、発根にエネルギーが回らなくなり、発根率が低下する。従って、挿し穂の芽は生長開始前の状態にあるものが良い。すなわち、このような芽の生長が盛んに行わないような芽をもつ挿し穂を選択する必要があり、挿し穂の採取時期や挿し穂の部位の選定などが必要となる。
①頂芽挿し(天挿し)を行う場合には、頂芽の生長期には実施しない。
②側芽を挿す場合(一芽挿しあるいは管挿し)、腋芽が萌芽していないものを用いる。
●穂木の養分蓄積
発根前の穂木は、切口からわずかに養分が吸収されるものの、基本的には茎内に含まれる養分でまかなわれる。また炭水化物についても、光合成は行われるものの、穂木採取前の葉に含まれている炭水化物に頼っている。従って、穂木採取前の親木の管理が極めて重要な要素となる。施肥との関係では、リン酸、カリの含量が高い親木から穂木を採取した場合には発根率が高くなることが知られている。
①挿し穂を採取する場合には、晴天が続いた日を選ぶ。
②親木の施肥は、窒素過多とせず、特にリン酸とカリウムを多めに与える。
③充分な潅水を行い、水ストレス(過乾燥と過湿)を与えない。
●挿し床の管理
挿し床の水分状態は、挿し穂の吸水能力と密接な関係がある。挿し穂の吸水能力は切口からの吸水に頼っているため、ある程度の加湿状態が必要となる。最も適した土壌水分状態は、総水分率(潅水直後の液相率)の50~60%程度の水分状態がよいとされており、pF値では1.5程度がよいと考えられる。この状態は、水分の補給だけではなく、発根過程の挿し穂への酸素の補給にとっても好適条件となる。挿し床の三層分布の中で、液相と気相がともに高い素材で、無菌的なものが適している。
①pF1.5程度の土壌水分(テンションメーターによる挿し床の管理)
②加水状態にしない(酸素補給のため)
③三相分布の液相と気相の割合が高い土壌の選択
④無菌の用土の選択
●病害の防除
挿し木における病害は3種類に分けられる。①
ピシウム、疫病、フザリウム、リゾクトニアなどの直接的な病害、②
維管束が菌により塞がれて枯死する、③ 線虫による食害
病害の進入は細胞壁の厚さと密接な関係があり、細胞壁が厚いものほど進入しにくくなる。一般に窒素過多の条件で栽培したものや若い枝は細胞壁が肥厚しておらず、薄いため、病害の進入を受けやすい。
①無菌の用土の選択
②雑菌の混入していない用水の利用
③未熟な穂木を用いない
④窒素過多で親木を管理しない
●温度管理
発根能力は25℃までは高まり、それ以上では低下する。これに対して病菌は30~35℃まで増殖率が高まることから、地温を25℃以上の温度としない管理が必要となる。
気温の上昇は蒸散を促進するため、気温を高めない。
①地温を高く維持する(25℃以上とはしない)
②気温を低く維持する(電熱温床の利用)
●発根阻害物質の除去
作物や品種によっては発根の困難なものがあるが、その原因の一つに発根阻害物質や褐変物質がある。これらの阻害物質の存在が認められる場合には、流水による浸漬処理が効果的である。
①発根阻害物質の存在がみられる場合には、流水に浸漬処理する
2)2)接ぎ木繁殖
接ぎ木(grafting)は、果樹の技術として発達し、中国では柑橘の接ぎ木が行われており、ヨーロッパでもギリシャ時代(紀元前300年)には既に実用的な技術として用いられていた。日本へは仏教の伝来とともに中国から伝えられた。現在栽培されている果樹のほとんどは接ぎ木繁殖によって増殖されている。
野菜の接ぎ木は1920年代から研究が進み、スイカなどのウリ科植物やトマトなどのナス科植物で行われている。
★台木と穂木
接ぎ木は根を持つ個体(台木:rootsrock)に芽を持つ枝(穂木:scion)を接ぐことである。台木と穂木を接ぎ木する際に重要な点は、両者の形成層を接合させることである。
1)接ぎ木の意義
①遺伝的に固定されていない品種の維持・増殖
果樹や花木では交雑によって育種が行われているが、これらの品種は種子で繁殖すると形質が固定しないため、栄養繁殖法の一つとして用いられる。
②突然変異個体の増殖
花木などでは突然変異による花色の変異などが品種開発に重要な役割を担っており(枝変わり)、これらを維持増殖する目的で接ぎ木が用いられる。ボタンはほとんどがシャクヤクに接ぎ木される。
③土壌伝染性病害虫の対策
土壌伝染性病害に対して抵抗性を示す品種を台木として用いる。特に野菜で利用が多く、スイカ93%、キュウリ72%、ナス50%、露地メロン44%、トマト32%などで台木が使用されている。国内で使用される野菜の台木は、年間5~6億本といわれている。
④不良環境耐性の付与
耐寒性、耐暑性、耐乾性、耐湿性の高い台木を使用することで、穂木の品種が適さない環境でも栽培が可能となる。例として、柿は亜熱帯果樹にも分類され、耐寒性が低く、低温での根の伸長性が低い。そこで、日本自生のマメガキに接ぎ木することで寒冷地での栽培が可能となる。同様にキュウリでは、冬季の栽培では低温での根の伸長性の高い"クロダネカボチャ"を使用し、夏季の栽培では高温耐性の強い"新土佐"を使用する。
⑤樹勢のコントロール
リンゴでは「わい性台木」といわれる台木を使用することで樹高が低く抑えられる。わい性台木による樹のわい化の機構は不明な点もあるが、根で作られる植物ホルモン活性が低いことや穂木と台木の物質伝達が不十分であるなどの原因が考えられている。野菜ではユウガオやカボチャにスイカを接ぎ木することで、樹勢が強く維持され、栽培期間の延長効果が期待できる。
⑥収穫物の品質向上
キュウリでは、通常の台木に接ぎ木するとブルーム(果粉)と呼ばれる白い粉が果実に付く。ブルームレス台木(カボチャ台木の一種)に接ぎ木するとブルームが発生しなくなり、光沢のあるキュウリが収穫できる。また、スイカをカボチャ台木に接ぎ木すると果実の肩が盛り上がり。カボチャ臭くなるが、ユウガオを台木にすると糖度が高くなる。またバラではノイバラに接ぎ木すると切り花本数が多くなり、花色が鮮やかになる。
⑦花芽分化の促進
通常接ぎ木をすると、台木と穂木との間の接ぎ木部位の維管束の結合が必ずしも順調にいくとは限らず、養分の移行が不良となることが多い。その結果、光合成産物が地下部に移行しにくくなり、接ぎ木上部に炭水化物が蓄積されるようになる。植物の花芽分化は、いわゆるC-N比が高まることによって促進されるといわれており、接ぎ木上部の炭水化物(C)の含量が高まることで花芽分化が促進される。「桃、栗3年、柿8年」といわれるように、柿は実生では8年程度の幼木期を経た後花芽分化するが、、マメガキや実生台木に接ぎ木をすることで2~3年で結実するようになる。
(2)接ぎ木の種類
接ぎ木法として、芽接ぎ、枝接ぎ、高接ぎ、腹接ぎ、切り接ぎ、根接ぎ、割り接ぎ、合わせ接ぎ、寄せ接ぎ、挿し接ぎ、接ぎ挿しなどがある。
3)株分け・分球(Division)
宿根草や球根植物では株分けや分球で繁殖される。一般に生きた根を付けた状態で分割する場合を「株分け」といい、根がない場合や根が死んでいる状態のものを分割する場合を「分球」という。球根植物の鱗茎では、分球のほかに鱗片挿しといわれる方法で繁殖する。